

RESEARCH
「環境の時」
原始の生命体と地球の姿
原始の生命体がくらしていた頃の地球環境に近いとされる海底熱水地帯にどんな生きものがくらしているのか。全ての生きものの共通祖先がもっていた遺伝子はどんな環境でくらすのに適していたのか。原始の生命がくらしていた環境を実験室の中につくるなど、進化を実験によって検証することで私たち全生物の共通の祖先の姿とくらしを浮かび上がらせたい。
1.有機物の合成から生命の誕生へ
今から約46億年前、地球は太陽系の他の惑星と共に誕生した。最初の地球は、隕石の衝突によるエネルギーのためにマグマのようなどろどろの状態で、隕石に含まれていた揮発性成分が蒸発してできた原始大気で覆われていたが、宇宙への熱の放出で急速に冷却していった。大気中の水蒸気は液体の水になり、原始大洋ができたと思われる。地球上で最古の岩石(約40億年前)には、すでに海が存在していた跡が残されている。
1953年に発表されたミラーの有機物合成実験(註1)以後のさまざまな研究により、アミノ酸など生体を構成する有機化合物が地球上で自然に生じる可能性があることがわかった。宇宙空間でも温度の低い場所では、宇宙塵の周りにできた氷の中で多くの有機化合物ができ、それが地球へと降り注いだ可能性もある。最初の熱い時期は分解してしまっただろうが、温度が下ってからは蓄積しただろう。
有機物は何億年もかけることで原始大洋に蓄積したと思われるが、生成した有機化合物が分解することもあり、どのような化合物がどの程度蓄積したのかは未だにはっきりとしていない。アミノ酸や有機酸は比較的簡単に生成するのに対し、核酸合成の鍵となるリボースが十分量蓄積したかは不明で、RNAが生きものの代謝によらずできたかどうかはわかっていない。
一旦RNAができれば、リボザイムとしてさまざまな反応を触媒するため、RNAが今はDNAが担っている遺伝情報も、またタンパク質が担っている触媒作用も担う世界(RNAワールド)があったと分子生物学者の間では当然の事実として信じられている。ただし、RNAが生きものによらず合成されたかは明らかでなく、RNAワールドは非常に危うい基盤の上に成り立っている考えであることも確かだ。
(註1) ミラーの有機物合成実験
1951年に大学院生であったミラーが当時考えられていた原始地球の大気(メタン、アンモニア、水素、水)をフラスコに入れ、放電(雷のモデル)したところアミノ酸などが容易にできた実験のこと。原始の地球環境で生きものを形づくる物質が生成することを再現した実験である。
2.化学合成生物の誕生
いずれにせよ最初の生命体は有機化合物のスープの中に誕生し、まわりの有機化合物を取り込んで増殖した従属栄養生物(註2)だったはずである。そこでは、複雑な有機化合物が次第に消費されるので、単純な化合物から複雑な有機化合物を合成できる生物が生き残りに優位になり、原始の生物に代謝系が発達していった。しかし、有機化合物がすべて消費しつくされれば、せっかく代謝系を発達させた生命も絶滅の危機に瀕する。そうなる前に無機化合物の自由エネルギーを利用して化学合成(註3)をする生物が誕生したはずだ。
(註2) 従属栄養生物
動物のように、自ら有機化合物を作り出すことができず、体外から取り込む生きもののこと。一方、植物などは自ら有機化合物を作り出すために独立栄養生物と呼ばれる。
(註3) 化学合成
有機物をつくるときに無機物の酸化還元エネルギーを利用する方法のこと。植物の場合は光エネルギーを利用するために光合成とよばれる。
3.化学合成細菌が支える海底熱水地帯の生態系
現在の生きものは、そのほとんどが太陽光のエネルギー(厳密には自由エネルギー)による光合成に依存して生きている。ところが近年、深海底の熱水噴出口周辺に地球のもつエネルギー(自由エネルギー)を利用する生きものたちの豊かな生態系がみつかった。もちろん、1000メートルを超える深海底に光は届かず、海水表面近くで行われた光合成による有機化合物やそれらを食べた動物の死骸も、海底へ沈殿する前に完全に分解してしまう。それにもかかわらず、深海底のなかでもチューブワームという名の1メートルを越える管状の生物や、多数の貝類、エビやカニ類などさまざまな生きものがくらしていたのだ。この思いがけない生態系は地上の火山と同じく地熱活動が活発な地域でみられる。地底から湧出する熱水中には海水が高温の岩石と反応して生じる硫化水素などの還元型の化合物が含まれている。
海底にはこの還元型化合物と海水中の酸素(あるいは酸化型化合物)の反応によって得られる自由エネルギーを利用して有機化合物をつくりだす化学合成細菌が存在し、この有機化合物に頼って前出したさまざまな生きものがくらしているのである。還元型化合物は地底深くで海水が高温の岩石と反応してつくられるのであるから、この生態系はいわば地球のエネルギーに依存した生態系と言うことができる。光合成が生み出される前の時代、地中から吹き出す熱水は唯一の大量な自由エネルギー供給源であった。そして、地球ができてまだ間もない時期には、地球内部の温度もまだ高く、現在よりも多くの海底熱水活動があったはずで、生命誕生とその後の生きものの世界は海底熱水地帯にあった可能性が高い。
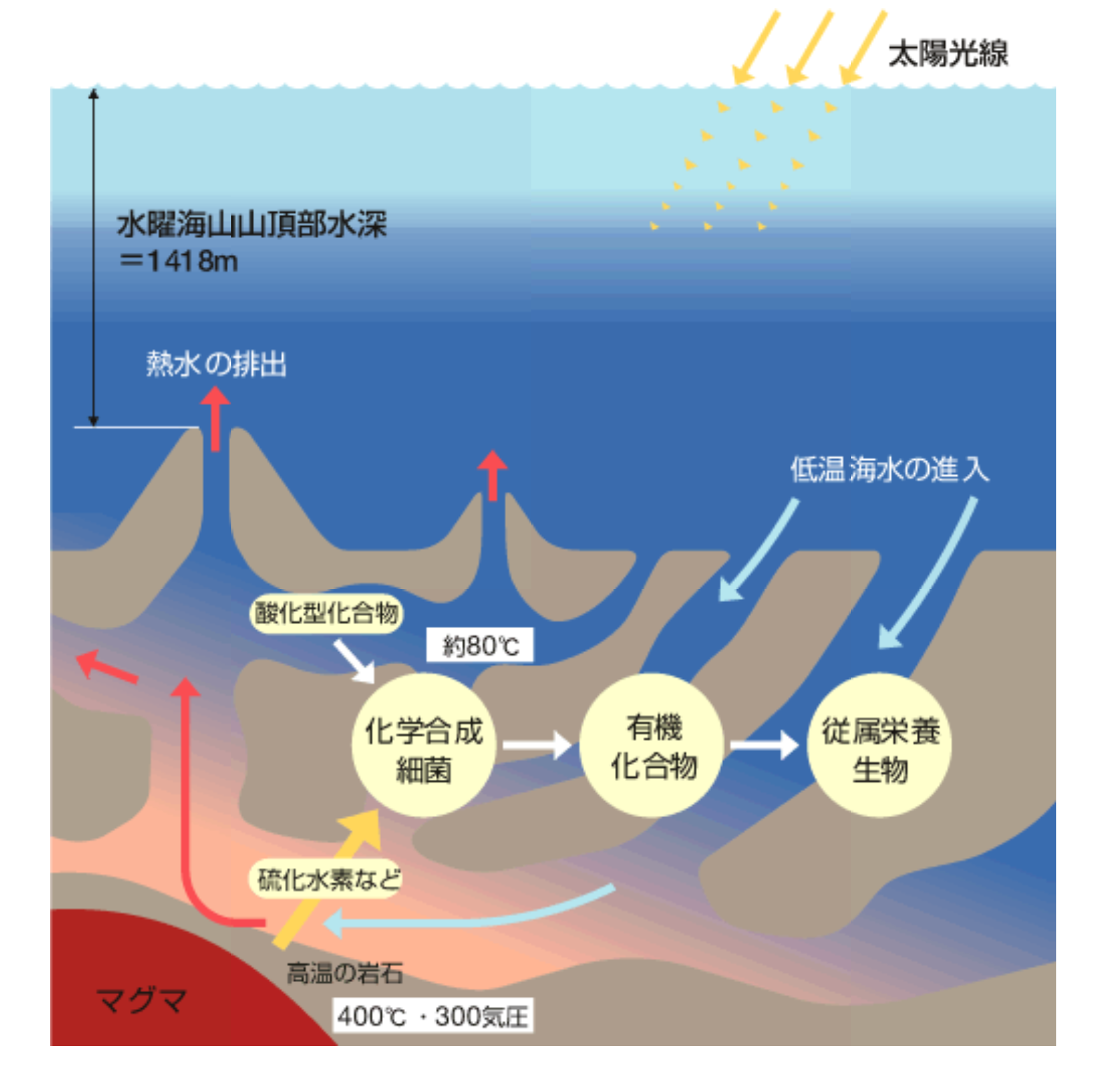
(図1)海底熱水噴出口付近
4.残された原始環境としての海底熱水地帯
つまり、現在の海底熱水環境は地球形成初期の原始の生物が暮らしていた環境を残しているといってよい。我々は現在、全国の大学と国立研究所に所属する地球物理学者、地質学者、地球化学者、微生物学者と共同で海底熱水地帯を解明する研究計画「アーキアンパーク」を進めている。まず、太平洋の真ん中(小笠原諸島の西より)にある水曜海山という海底火山の火口に特殊な海底掘削装置で穴を掘り、火口地下の構造とそこに棲む微生物の分析を行った。
火口地下にどんな生きものがくらしているかを知るために、掘削孔から採集した熱水に含まれるDNA中のリボソーマルRNA遺伝子(註4)配列を解析し、その中の生物種を推定することにした。熱水中の生きものの大半はまだ人間が培養したことのない新規のものであったが、DNA配列を既知の生きものと比べることで 何に似ているのかは推定できる。その結果、深海の熱水中の水素や硫化水素を利用している化学合成細菌に似ていることがわかった。
(図2)水曜海山の位置
もちろん、DNA配列が似ているからといってそれらが実際に水素や硫化水素を利用できる化学合成細菌であるという保証はなく、培養して確かめる必要がある。しかし、どのような微生物かわからなければ培養できない、培養できなければどのような微生物かわからないという袋小路に入ってしまう。そこで、そこから抜け出すために、実験室内に海底熱水系の環境を作ろうと考え、いまその計画を進めている。装置内は400度、300気圧という超臨界条件に達し、つくりだされた海底熱水が低温の海水中へと噴出するように設計されている。この装置に海底熱水地帯から採集したサンプルを入れれば、これまで培養されたことのない微生物が増殖するはずだ。それを期待している。
(註4) リボソーマルRNA遺伝子
リボソーマルRNA遺伝子は細胞内でタンパク質を合成するというとても重要な機能をもち、全ての生きものがこの遺伝子(機能も同じ)を持っている。そのためこの遺伝子の配列を比較することで、生きものの系統解析ができる。そのうえ、重要な機能をもつことから遺伝子配列が進化する速さが遅く、古い時代に分かれた生きものの系統を解析するのに適している。
5.遺伝子から見えてきた全生物の祖先のくらしていた環境
原始の生きものとそれがくらしていた地球環境を知るためのもう一つのアプローチとして、当時の生きものの持っていた酵素(タンパク質)を実験的に再現しようという研究も進めている。
現存生物はすべてその祖先の遺伝子を受け継いでいるので、さまざまな生物の遺伝子を比べて進化系統樹を作ることができる。すべての生きものを比べる時によく用いられるのが、先ほど述べたリボソーマルRNA遺伝子である。生物にとって不可欠であり、基本は変わらないまま進化をしてきた遺伝子であるために、比較に適しているからだ。リボソーマルRNA遺伝子による系統樹(図3)では大きく真正細菌、古細菌、真核生物の3グループが分かれ、興味深いことに生育温度の高い生物がその根元に集まる。つまり、全生物の共通な祖先(コモノート)に近いと思われる生きものに高温を好むものが多いのである。
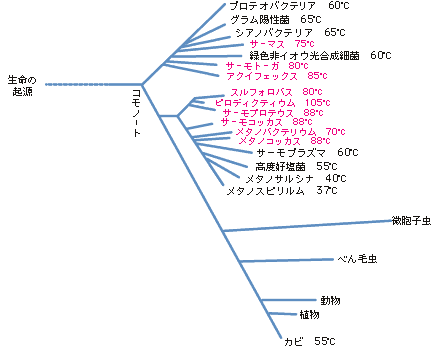
(図3) リボソーマルRNA遺伝子によってつくられた全生物の進化系統樹(Woeseらによる)
数字はその生物群の中で最も生育温度の高い生きものの生育温度。特に70℃以上の場合には赤で示してある。
本当にコモノートは高温を好んだのだろうか。我々は特定の酵素を選んで、系統樹を作ってみた。実際には全生物が持っており、研究室で扱い慣れているイソプロピルリンゴ酸脱水素酵素(IPMDH)の遺伝子を用いた。さらに、系統樹を作るときに用いた各生物のIPMDHのアミノ酸配列を比較し、そこから共通祖先の持っていたIPMDHのアミノ酸配列を推定したのである。次に系統樹で最も祖先に近いとされた超好熱性古細菌Sulfolobus tokodaii(註5)のIPMDH(今まで調べられたIPMDHの中で最も耐熱性が高い)を土台にして、推定した共通の祖先型のIPMDH配列に近づくようアミノ酸配列を部分的に変えたIPMDHを7種つくった。そして、それらの耐熱性を調べてみると、そのうちの5つは元のSulfolobus tokodaii のIPMDHよりもさらに耐熱性が上がった。
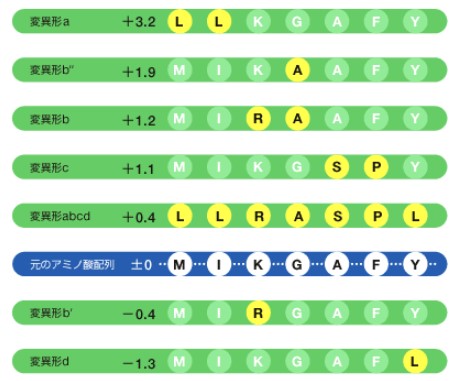
(図4)祖先型の配列に近づけたIPMDHの耐熱性の変化(℃)
Sulfolobus tokodaiiのIPMDHアミノ酸配列の中からターゲットとなるアミノ酸(白丸)を決め、それを祖先型のアミノ酸(黄丸)に変えたIPMDHをつくり、耐熱性の変化を調べたところ、7つ中5つで耐熱性が上がった。数値は、もともとのSulfolobus tokodaiiのIPMDH耐熱性に比べて変化した温度(℃)。耐熱性の大きい順に並べてある。
A=アラニン、G=グリシン、F=フェニルアラニン、I=イソロイシン、K=リジン、L=ロイシン、M=メチオニン、P=プロリン、R=アルギニン、S=セリン、Y=チロシン
これは、全生物の共通の祖先は現在の超好熱菌よりもさらに高温で生育していたことを示唆しているのではないだろうか。現在、すべての遺伝子の祖先型遺伝子を作成する計画を立てており、いつか、祖先型の生物をなんとかよみがえらせたいと思っている。
(註5) Sulfolobus tokodaii
大分県別府温泉から採取された古細菌。75~81℃で酸性(pH3)という過酷な環境を好んで生育する。
山岸明彦(やまぎし・あきひこ)
1981年東京大学大学院理学系研究科博士課程修了 、理学博士。
日本学術振興会奨励研究員 、東京大学教養学部の助手、カリフォルニア大学バークレー校、
カーネギー研究所の博士研究員 、東京工業大学理学部(現生命理工学部)助手を経て
現在東京薬科大学生命科学部助教授 。
.jpg)

.jpg)
.jpg)










