

RESEARCH
新天地を目指して
―陸上への引っ越しと器官のリサイクル
最近、長らくお世話になった国立遺伝学研究所(静岡県三島市)を離れ、東京に移った。不要になった芝刈り機や自転車、布団干し、バーベキューセットなどを処分し、新天地での生活に必要なものを新たに手にいれた。新しいものを購入するのもお金がかかるが、不要なものを処分するのも負担が大きい。いらなくなったものから必要になったものをつくれたらよいのにと真剣に考えた。
まずはここから…リサーチのツボ
研究マップ:陸上への引っ越しと器官のリサイクル
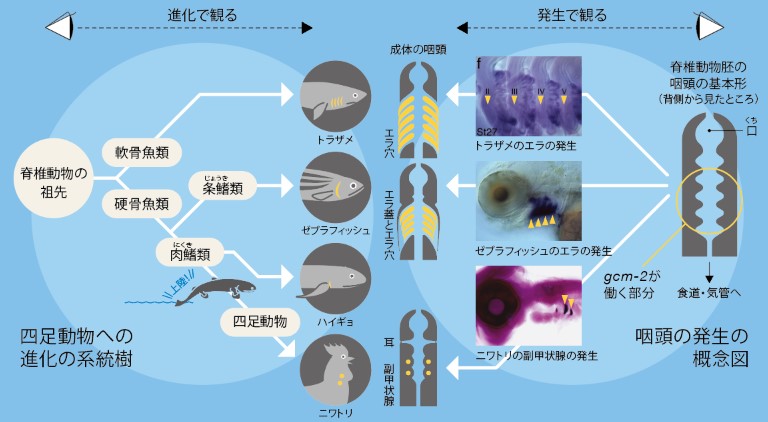
1.上陸への必需品-副甲状腺
今から約3億7千万年前、我々の祖先は住み慣れた海から陸に上がった。この「水中から陸上への引っ越し」は、乾燥や重力などすぐ思いつくものも含めて想像を絶する環境変化であり、不要になったものと新たに必要になったものとが山のようにあったに違いない。それは、陸上に住む脊椎動物(四足動物)の外見が水中に住む脊椎動物(魚類)のそれと大きく異なっていることからもわかる。環境の変化に合わせて変化したのは外見だけではない。血中の酸素濃度や体液のイオン組成など、細胞が働くための体内環境は外界の変化によらず、一定に保たれなければならない。その工夫の一つが、四足動物に見られる副甲状腺である(図1)。
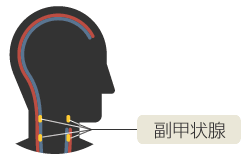
(図1)首の中にある副甲状腺
米粒状の器官で、左右に2つずつ合計4つ存在する。
副甲状腺は、血液中のカルシウムイオン濃度を一定に保つための内分泌器官である。細胞外(血液中)のカルシウムは、筋肉の収縮、神経刺激の伝達、血液の凝固、ホルモンの分泌など、多くの重要な生理機能を調節している。特に細胞外から細胞内へのカルシウムの流入は、多くの現象のシグナルとして働くので、血液中のカルシウム濃度は一定に保たれていなければならない。四足動物はカルシウムを食餌からしか得ることができないので、骨にカルシウムを蓄積し、濃度が低下する度に血液中に適量溶かし出す必要がある。副甲状腺は、血液中のカルシウム濃度を監視し、副甲状腺ホルモンを血液中に分泌して全身の骨からカルシウムを溶かし出し、血液中のカルシウム濃度を一定に保つ司令塔なのである。
一方魚類は、血液中のカルシウム濃度が低下した場合には、周囲の水からカルシウムを取り込めるのでわざわざ骨にカルシウムを備蓄する必要がない。むしろ魚類は高濃度のカルシウムを含む水に常に晒されているので、血液中のカルシウム濃度を低下させるのに躍起になっているというのが実情である。つまり新天地を目指した我々の祖先は、突如、副甲状腺を生活必需品として手に入れなければならなくなったのである。ご先祖様はいったいどうやってこれを入手したのだろう。
進化の道筋を知るためには、化石を調べるのが手っ取り早い。しかし内蔵や筋肉といった柔らかい組織に関する情報を、化石だけに求めても十分なものは得られない。この疑問を解くもう一つの方法は、現生生物のいくつかで、上陸と密接に関連すると思われる器官がどのような発生プロセスを経て生じるのかを明らかにし、それを比べることである。この方法で、その器官が本来はどのような構造を持っていたものか、またどのように進化したのかを推測できるに違いない。
2.咽頭に生じる様々な器官
脊椎動物の上陸時に、首の中に副甲状腺が現れた。一方消えていったのは、肺呼吸で不要になったエラである。四足動物はエラを持たないが、発生過程には魚類のエラとよく似た構造「咽頭嚢(いんとうのう)」が生じる(図2)。咽頭上皮(内胚葉性上皮)が外側方に延び、袋状の構造を前後に約5対形成するのだ。魚類の場合、この咽頭嚢が外胚葉上皮と融合してその部分に穴が開き、エラ穴を作る。四足動物ではこれらの咽頭嚢はエラ穴にならず、その窪みから様々な構造物や器官が発生する。例えばヒトの場合、1番前の咽頭嚢(第1咽頭嚢)は耳管(エウスタキオ管)と中耳鼓室腔(鼓膜の裏の耳小骨が存在する空間)とになる。2番目の咽頭嚢は口蓋扁桃(俗にいう扁桃腺)を、3番目の咽頭嚢は胸腺(免疫細胞の分化に関わる)と副甲状腺を、4番目は副甲状腺のみを産み出し、一番後ろは甲状腺の傍濾胞細胞(カルシトニン産生細胞)を産生する。
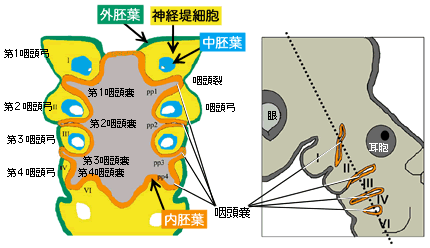
(図2) 咽頭嚢の模式図
左:ニワトリ初期胚の頸部の内部構造。右の図の胚で、破線の領域を切断面として見たもの。
脊椎動物の発生過程において、頭部側の消化管に左右数対の膨らみが生じる。この領域は将来の口腔や食道に対応するので、咽頭嚢の名が冠せられている。ニワトリでは第1から第4まで4つの明瞭な咽頭嚢が形成され、5番目は発生せず6番目も痕跡的である。なお、脊椎動物胚の頸部にスリット状の構造(咽頭裂)が見えるのは、咽頭嚢の内胚葉に接している外胚葉が内側に落ち込んでいるため。前後の咽頭嚢や咽頭裂に挟まれた領域は、中胚葉細胞と神経堤細胞が外胚葉上皮と咽頭嚢内胚葉に囲まれた咽頭弓(いんとうきゅう)と呼ばれる構造になる。咽頭弓からも、アゴや耳小骨など様々な器官が発生する。
興味深いことに、どの咽頭嚢から何ができるかは、動物によって多少異なっている。例えばニワトリでは、ヒトとは異なり胸腺が第3咽頭嚢だけでなく第4咽頭嚢からも発生する。一方マウスの副甲状腺は第3咽頭嚢からしか発生しないのに対し、トカゲでは第2咽頭嚢からも副甲状腺が発生する。一番後ろの咽頭嚢から発生するカルシトニン産生細胞は、哺乳類では甲状腺の中に移動するが、それ以外の四足動物では甲状腺の中には入らず、鰓後体という独自の内分泌器官を形成する。
このように、種によって免疫系組織(扁桃や胸腺)と副甲状腺を発生させる咽頭嚢が異なるということは、元来第2~4咽頭嚢はこの2種類の器官を発生させるポテンシャルを持っており、進化の過程で各動物が都合のよい咽頭嚢からそれぞれを発生させるようになったということかもしれない。それを確かめるには、より原始的な脊椎動物の発生を調べればよいわけだが、魚類の咽頭嚢からは胸腺は発生するものの、副甲状腺は生じないので実は簡単ではない。四足動物に副甲状腺が存在する理由を知るためには、魚類に副甲状腺が存在しない理由も知らなければならないことになってきた。しかし、存在しない器官の発生を調べるにはどうすればよいのだろう。
3.エラと副甲状腺-転用された遺伝子の働き
生物はそれぞれが持つゲノム情報に基づいて自らの体を発生させる。その結果が我々の目の前にいる多様な形態を持つ生物たちだ。生物種ごとに異なる形態が生じるのは、進化の過程で蓄積した様々な遺伝子の変異が、発生プロセスに変更を加えていったためと考えられる。解剖学的に魚類には副甲状腺がないわけだが、それは器官としての細胞塊が見つからないというだけかもしれない。つまり、魚類のゲノムの中にも副甲状腺の発生に関わる遺伝子は存在するけれど、四足動物とは使い方が異なっているという可能性はないだろうか。問いをこのような形に変えてみた。哺乳類で、咽頭曩から副甲状腺が形成される過程は、Gcm-2という遺伝子によって制御されていることがわかっている。我々はニワトリでもGcm-2が副甲状腺に特異的に発現していることを明らかにした(図3)。そこで、この遺伝子を副甲状腺のマーカーとして使い、魚類に副甲状腺の起源を探すことにした。
オーストラリアハイギョ、フグ、ニジマス、ゼブラフィッシュ、トラザメといった各種魚類からからGcm-2の相同遺伝子をクローニングした。その結果、副甲状腺を持たない硬骨魚類や軟骨魚類にもGcm-2類似の遺伝子が存在することがわかった。分子系統樹を作成してみると、これらの魚類のGcm-2様の遺伝子はいずれも、他のGcmファミリーに属する遺伝子よりも、四足動物のGcm-2に似ていた(図3)。さらにヒト、マウス、ニワトリ、ゼブラフィッシュ、それぞれのGcm-2遺伝子座付近のゲノム構造を調べると、どの遺伝子でも隣にある遺伝子が同じであることがわかった。これは、クローニングした魚類のGcm-2様の遺伝子は四足動物のGcm-2と同じ遺伝子であることを示している。
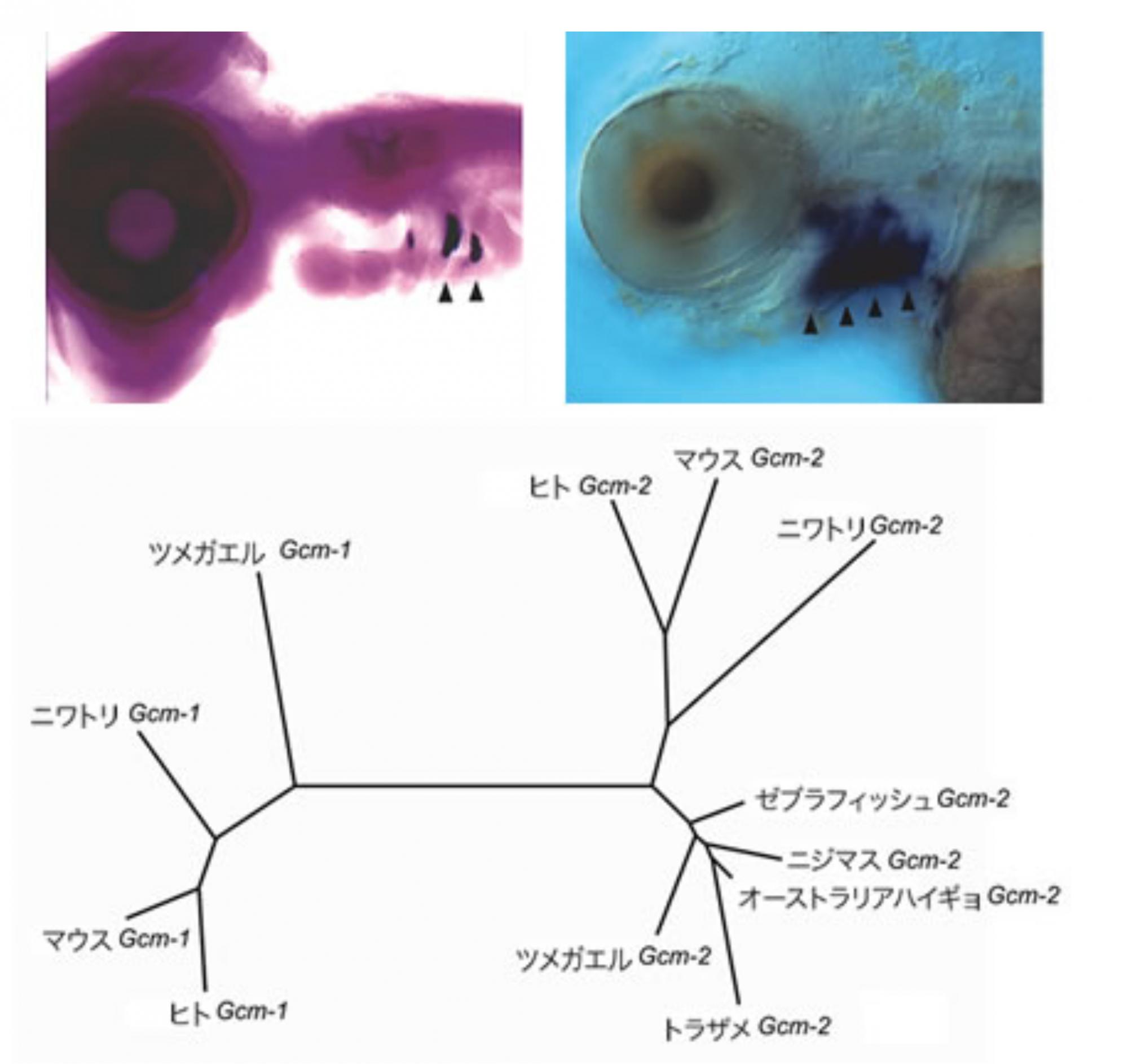
(図3) 咽頭嚢の発生とGcm-2遺伝子
左上:ニワトリ胚で発現するGcm-2(矢頭)。この領域は将来副甲状腺となる。右上:ゼブラフィッシュ胚では、将来エラとなる領域でGcm-2が発現していた(矢頭)。下:脊椎動物の系統とGcm遺伝子ファミリーの系統図。枝の長さが長いほど、お互いの遺伝子配列に違いが多いことを示す。Gcmファミリー遺伝子にはGcm-1とGcm-2の2つが存在するが、エラや副甲状腺で発現するGcm-2は全て同じグループに属している。
硬骨魚類ゼブラフィッシュにおけるGcm-2の発現パターンを調べてみると、発生中の鰓芽(エラの原基)に強く発現していることがわかった(図3)。鰓芽は副甲状腺と同じように咽頭嚢の内胚葉上皮から発生し、将来エラになる。さらにアンチセンスモルフォリーノ(※註)を用いてゼブラフィッシュ胚におけるGcm-2の機能を抑制したところ、鰓芽が発生せず、魚類においてGcm-2はエラの形成に必要であることがわかった(図4)。
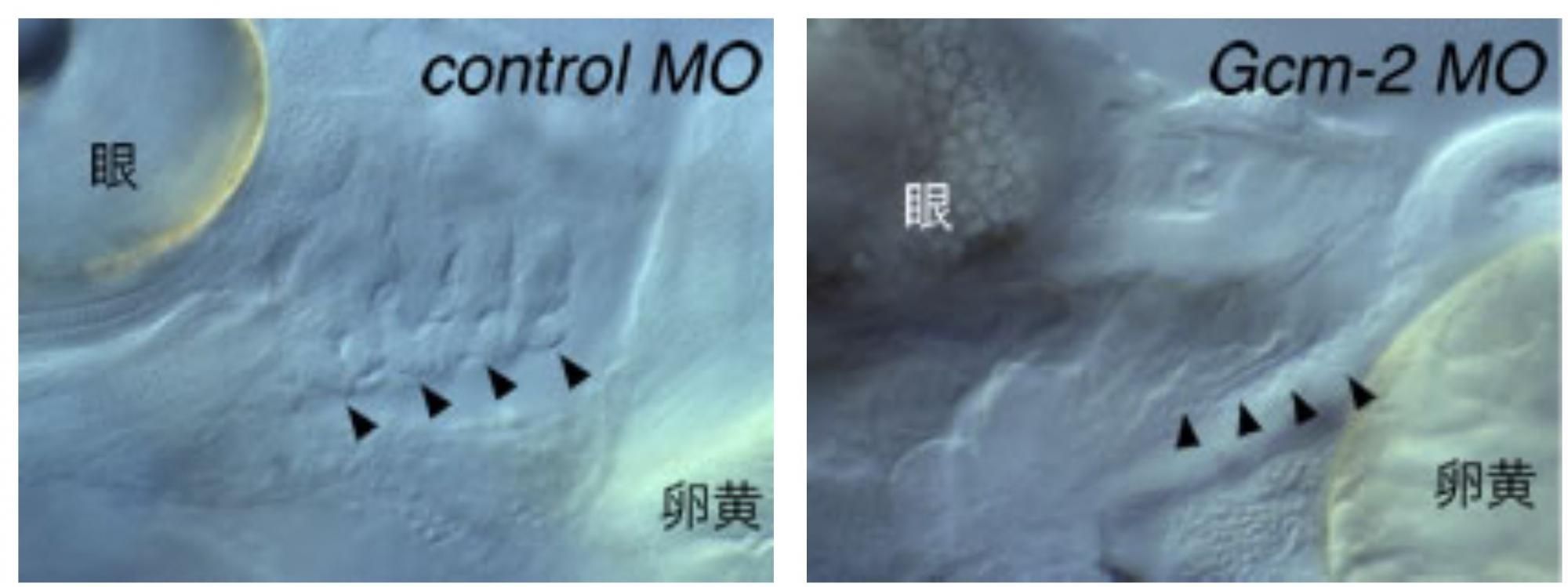
(図4)Gcm-2が働かないとエラができない
左:正常胚では鰓芽が形成され、これが将来エラとなる(矢頭)。右:Gcm-2の機能を阻害すると、鰓芽が形成されなくなった
このように硬骨魚類までは辿れたので、さらに進化の過程をさかのぼり、四足動物と硬骨魚類が分かれるよりはるか以前に共通祖先から分岐した軟骨魚類におけるGcm-2の発現パターンを見た。具体的にはトラザメ胚を調べたところ、ここでもGcm-2は鰓芽に発現していた(図5)。つまり、Gcm-2という遺伝子は元来エラの発生に必要な遺伝子であり、陸上に上る時にその機能を副甲状腺の発生に転用したと考えてよさそうである。
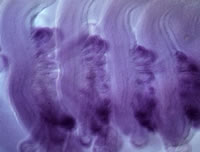
(図5) トラザメ胚の鰓芽に発現するGcm-2
(註)アンチセンスモルフォリーノ
モルフォリーノとは、核酸を構成するリボースと類似した性質を持つ物質の名称。デオキシリボースのかわりにモルフォリーノを使って合成したDNA類似物質(モルフォリーノオリゴ)は、核酸分解酵素などの作用を受けず非常に安定である。細胞が作るmRNAと相補的な配列を持つモルフォリーノオリゴを細胞内に導入すれば、モルフォリーノオリゴとmRNAが強く結合し、mRNAからの翻訳が阻害される。遺伝子組み換えなどゲノムを直接操作することなく、遺伝子の機能を阻害する方法として利用されている。
4.エラの意外な役割
エラの細胞が副甲状腺に変化したのなら、発生を司る遺伝子の共通性だけでなく、生理機能的な類似性も存在するかもしれない。魚類では周囲の水に溶けているカルシウムイオンが体内に入るが、その内約70%がエラを介して取り込まれている。副甲状腺は血液中のカルシウム濃度を監視し、副甲状腺ホルモンを分泌することがその役割であるが、エラにも同様の機能があるのではなかろうか。次に立てた問いはこれである。
これまで魚類には副甲状腺ホルモンはないとされていた。魚類では骨内のカルシウムを血液中に再吸収させる必要がないので、これは道理にかなっている。魚類には解剖学的に副甲状腺がないこともあり、あまり精力的に魚類に副甲状腺ホルモンを探すことはなされてこなかった。ホルモンそのものを物質として探すのは面倒な仕事だからである。しかしゲノム情報が集積した現在では、魚類ゲノム中に副甲状腺ホルモンを生成する遺伝子を探すことができる。これならホルモン探しより面倒が少ない。そこで、ゼブラフィッシュとフグのゲノム塩基配列を調べたところ、共に副甲状腺ホルモン遺伝子が2つずつ見つかった(図6)。さらにゼブラフィッシュゲノムから、副甲状腺が血液中のカルシウム濃度を感知するために用いているカルシウム感受性受容体の遺伝子を同定し、これらの遺伝子の発現をゼブラフィッシュ胚で調べてみた。2つの副甲状腺ホルモンとカルシウム感受性受容体はどれもエラで作られていることがわかった(図6)。発生学的にも、生理学的にも、エラと副甲状腺はよく似た存在であると考えることができ、副甲状腺がエラから進化してきた可能性を示す結果を得ることができた。
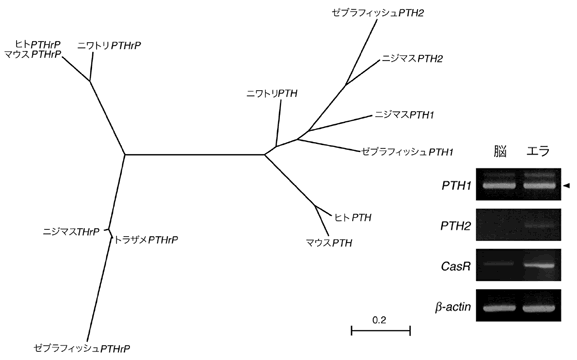
(図6)魚類は副甲状腺ホルモンを持ち、カルシウム濃度もエラで感知してい
左:副甲状腺ホルモン(PTH)の系統樹。副甲状腺ホルモン類似タンパク質(PTHrP)のみならず、副甲状腺ホルモンそのものを魚類は持っていた。
右: ゼブラフィッシュにおける2つのPTHとカルシウム感受性受容体遺伝子(CasR)の発現。成魚の脳とエラからRNAを抽出し、PTHとCasRの配列を持つRNAが存在しているかを調べた。2つのPTHとCasRはエラで発現しており、これらの遺伝子が、偽遺伝子(タンパク質をコードしていても発現しない“死んだ”遺伝子)ではないことが示された。なおβ-actin(ベータアクチン)はほぼ全ての組織で発現することがわかっており、この実験において操作の誤り(RNAの抽出ミスなど)がなかったことの証明として同時に検出したものである。
5.器官のリサイクルを読み解く
副甲状腺の役割と言えば、血液中のカルシウム濃度を監視し、血液中にホルモンを分泌することだけである。血管がありさえすれば「足の裏」でも「筋肉の中」でも「骨盤の中」でもその機能を十二分に果たすことができるはずである。しかし実際には首にある。これは副甲状腺が魚のエラの成り代わりとして咽頭嚢から発生したからだと考えてよかろう。ヒトを含む四足動物はエラを捨て副甲状腺を新たに獲得したのではなく、不要になった品を新天地での生活に必須な副甲状腺に作り替えて首の中で今も大事に使っているのである。
脊椎動物が上陸する際に生じた形態変化のうち、内蔵や筋肉といった化石として残りにくい軟部組織の変化を調べるには、今回我々が試みたように特定の遺伝子やゲノム情報を利用したアプローチを併用するのが有効であることがわかった。そこで、ゲノム情報が急速に蓄積されているのだから大いに活用すれば理解が進むと期待するのは当然である。しかし実際はそう楽観的ではない。生物の形の多様性が生じるカンブリア紀の2億年も前にすでにゲノム構造は多様化していたことが知られており、遺伝子の変化が直ちに形の変化に結びつくものではないとわかっているからだ。新しい形質の獲得は、既存の構造を少しずつ変え、新しい生活空間に適した構造を生み出すことにより達成される。多数の遺伝子ツールはすでにゲノムの中に準備されており、いくつもの可能性の中から多様な形態を作り上げるような使い方の変化が生じたと考える方があたっている。つまりゲノム情報だけでは新しい形態の成立を理解するのに十分でなく、異なる形態を持つ動物間での発生現象を比較し、鍵となる細胞群間の時間的空間的な関係を詳細に解析していくことが、最も重要であることにかわりはないのである。
新天地への引っ越しに際して、不要になったものをリサイクルし新しい生活に役立つものを作り上げてきた私たちのご先祖様。副甲状腺は、生きものがつながりと変化を基本に進化してきたことの大事な証拠なのである。生物の進化の基本がここにあるとしたら現代人の消費型のライフスタイルは、生きものとしての生き方からははずれているということになるのではなかろうか。
岡部正隆(おかべ・まさたか)
1996年東京慈恵会医科大学大学院医学研究科博士課程修了。科学技術振興事業団CREST研究員、国立遺伝学研究所助手、ロンドン大学キングスカレッジ客員講師を経て、2005年より東京慈恵会医科大学DNA医学研究所・器官発生研究室室長。
.jpg)

.jpg)
.jpg)









