

RESEARCH
減数分裂
多様さを生み出す厳格なしくみ
1.多様な生きものを生み出した有性生殖
38億年前に生まれた最初の細胞は分裂によって自分と全く同じゲノムをもつ細胞を生み出した。このように自分のコピーを残す無性生殖が現在も続いている一方、約10億年前に新たなしくみが生まれた。二つの細胞が融合しゲノムを混ぜ合わせて新しい個体を生み出す有性生殖である。ゲノムを混ぜ合わせると、親とは異なる個体が生まれる可能性が高まるので、現在の多様な生きものを生み出す原動力となったと考えられ、日常目に見える生きもののほとんどが有性生殖によって子孫を残している。人間もその一員であり、精子、卵に含まれるそれぞれ23本の父由来と母由来の染色体を受け継いで、46本の染色体をもつ受精卵として人生をスタートする。実は二つのゲノムの混ぜ合わせだけなら細菌でも見られ、有性生殖に限ったことではない。約10億年前に起こった有性生殖が生み出された時に新しく登場したのは、混ぜ合わせた染色体を正確に半分にして生殖細胞をつくる減数分裂のしくみであり、しかもその巧みさはよくぞこのようなしくみをつくったと感嘆させるものである。減数分裂のしくみは有性生殖を行う生きもの全てに、完全に保存されている。そこで、減数分裂の分子的な基盤の研究が重要と考え、取り組んだ。
2.自分のコピーを生み出す細胞分裂とゲノムを半分にする減数分裂
減数分裂に入る前に、さまざまな細胞分裂のしくみを見てみよう(図1)。単細胞真核生物や多細胞生物の体細胞の分裂ではI状(アルファベットのIの形)の染色体が複製され、接着した状態で細胞の真ん中に整列した後、動原体付近を除いて接着が外れていく。動原体は染色体の中心付近にあることが多いので、染色体はX状になる。真ん中に整列するのは細胞の両端から伸びる糸がX状の染色体の中心部分(動原体付近)に結合して力を加えているからである。この糸に両側からひっぱられた染色体は両極へと分かれていく。ここで、同じ方向から引っぱられると、糸が外れてしまう。複製された染色体が動原体で接着していることが、両側に引っぱられていることを感知するために必要なことなのである。染色体が無事分かれた後に、細胞質の分裂が起きて元と同じI状の染色体をもつ細胞が二つできる。原核細胞のゲノムDNAは環状DNAで、染色体構造はとらないが、複製されたDNAが二つの細胞へ受け渡され、自分のコピーができるという点は、体細胞分裂と全く同じである。これらは38億年前から続く、生きものが自分のコピーをつくる際の基本原理であり、同じゲノムが正確に引き継がれることを保証している。
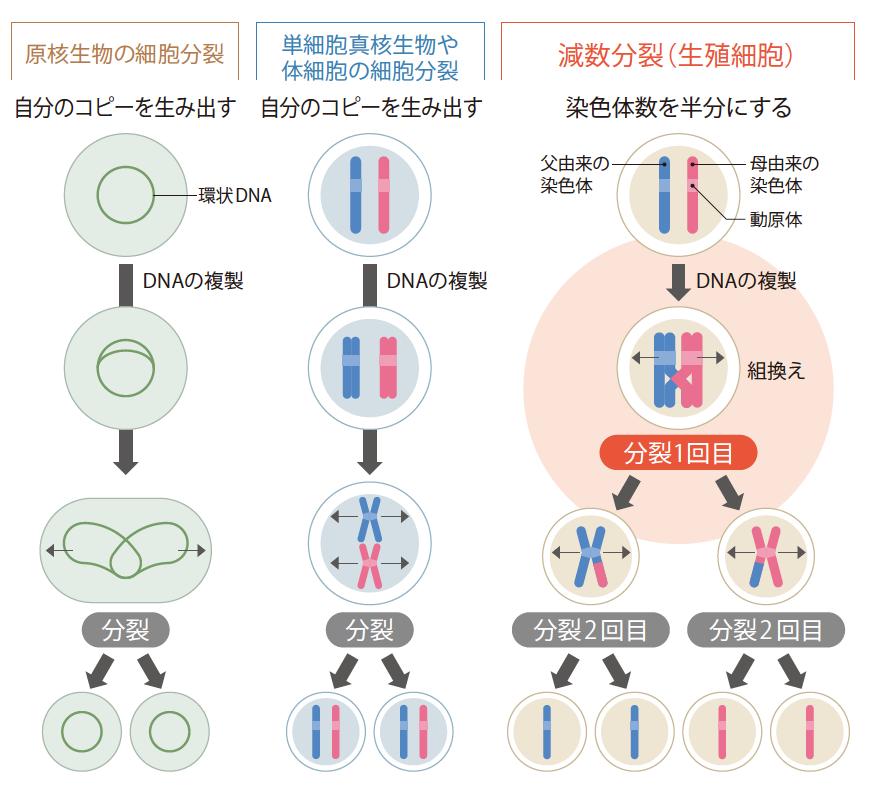
(図1) さまざまな細胞分裂の様式
次に減数分裂を見てみよう。減数分裂と呼ぶのは2回の分裂によって、染色体数を元の半分にするからである。かなり複雑なので図をよく見ながらしくみを追って欲しい。まず、複製された父由来、母由来の染色体が対になり、一部を組換えることでつながり、1回目の分裂にはいるが、その時にはそれぞれX状のまま別の細胞に分けられる。2回目の分裂はX状の染色体を均等に分け、それぞれI状になるという点で体細胞分裂と同じであるが、既に一度分裂しているので染色体数は半分になる。減数分裂の特徴は父由来と母由来の染色体が組み換えによってつながり、それぞれ複製された染色体がくっついたまま分かれる1回目の分裂にあると言える。
3.減数分裂を支える三つのしくみ
減数分裂を特徴づける1回目の分裂では他の細胞分裂とは異なる3つの性質が知られている。
つまり、
・ 父由来と母由来の染色体を組換える
・ 動原体付近の接着を守る
・ 複製した染色体を同じ方向に引っぱる
である。
実はこれらのしくみのそれぞれは有性生殖の誕生の時に発明されたものではない。組換えについては、体細胞では宇宙線などで傷ついたDNAは組換えに似たしくみで修復することが知られている。通常の細胞分裂でも複製した染色体は両方から引っぱられるのに耐えなければならないために動原体付近で強く接着しており、分裂の中期では染色体がX状に見える。また、体細胞分裂でも父親と母親の染色体を人工的につなげると、複製したX状の染色体がI状ではなく、X状のまま分けられてしまうことが知られている。そういう意味で、減数分裂の第一分裂の特徴といわれる3つのしくみは、既に体細胞の中にあったしくみの使い回しと言える。約10億年前にそれらが組織化され、減数分裂が誕生したのである。私たちはこの3つのしくみを詳しく調べた。
(ア)父由来と母由来の染色体を組み換える
「DNAの組換えは何のためにあるのか」と問われると多様化するためという言葉が第一に返ってくることが多い。確かに、組換えによって父由来と母由来の染色体が混ざり合うので多様化に貢献するだろう。しかし、組換えがなくとも、受精だけで充分に多様化はできる。例えば、ヒトの場合は染色体が23対、46本あり、いずれの染色体も減数分裂の際に2分の1の確率で選ばれるので、生まれる子どものもつ染色体の組み合わせは246 ≒ 約7兆通りもある。これだけあれば一人一人の違いを出すのに充分である。
ところで、父由来、母由来の染色体を組換える場所や数は生き物によって様々であるが、ヒトでは平均二箇所、マウスでは一箇所と意外と少ない。しかも、複数箇所ある場合でも、すぐ近くでは組換えが起きないように排除する機構があるので等間隔に組換えが起こる傾向がある。多様化しようとするなら多くの箇所で組換えた方がよかろうに、現実には組換えの数は少なく、むしろ一回だけの方がありがたいようにみえる。
私たちは組換えの本質を探るために、組換えを抑制する操作を施した酵母での減数分裂の分裂1回目の様子を観察した(図2)。すると、通常通り父由来、母由来の染色体が正しく両極へ移動する場合と、誤って一つの極に偏る場合が半々になった。父由来、母由来の染色体の引っぱられる向きが完全にランダムになってしまうのである。この結果から、組換えは父由来、母由来の染色体を正しい方向へ分配するために必要なしくみであり、そのために行われるものと考えてよいだろう。
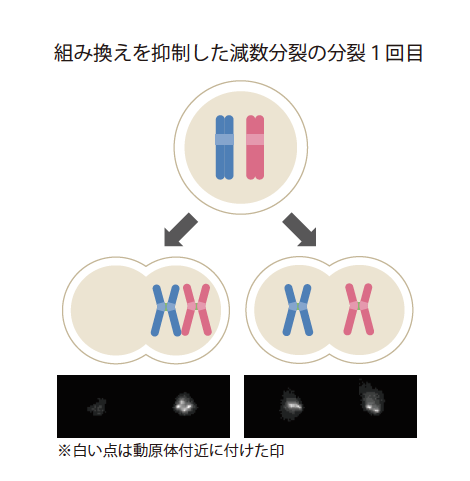
(図2) 染色体の正確な分配に大切な組換え
酵母の正常な減数分裂の分裂1回目では、組換え後に父由来と母由来の染色体が二つの細胞に分配されるのに対し、組み換えを抑制すると、父由来と母由来の染色体が二つの細胞に分配されることもあれば、片方に偏る事もある。どちらになるかは完全にランダムとなる。
減数分裂の分裂1回目では複製された染色体が体細胞分裂同様に接着するのに加え、組換えによって父、母由来の染色体もつながる。このため父由来と母由来の染色体は細胞の両端から伸びる糸によって反対向きに引っぱられる力を感知できる。組換えを起こさないように操作を施すと、父由来、母由来の染色体が分裂前に離れてしまい、反対向きに引っぱることができない。通常、全ての染色体が反対向きに引っぱられていることが確認されてから、染色体は両極に運ばれるので、組換えを起こさない場合は、そこで一時停止してしまう。酵母の場合には、20分ほど待っても反対側から引っぱられる力を確認できない場合には、間違っていてもいいやと思っているかのように、染色体をランダムに両極へ分配してしまい、異常分裂で死んでしまう。
私たちの結果は、組換えが多様化のためだけでなく、染色体の正しい分配に重要であることを示した。組換えによってつながることが正確な分配に役立つというのは、パラドキシカルではあるが、いやそれだからこそ非常に興味深い。
(イ)動原体付近の染色体の接着を守る
顕微鏡下で細胞分裂中の染色体を観察するとX状の形になっていることは、何十年も前から知られてきたが、その形を制御しているしくみは全く分かっていなかった。私たちはその一端を担う役者として、減数分裂の際に複製された染色体を接着する糊分子(Rec8)を99年に発見した。減数分裂の際に複製によって2本になった染色体の全体はRec8によって接着する。その後、父由来と母由来の染色体の間で組換えが起こる時も、複製で2本になった染色体間の接着は維持されたままである。染色体が細胞の両極から引っぱられる段階になって初めてRec8がはずれるのである。ただし、この時動原体の近くだけはRec8が残り、接着したままであることが重要である。父由来と母由来のX状の染色体がそれぞれ細胞の両極へと運ばれ、続いて2回目の分裂が始まるのだが、糊をはずす分子の働きは一回目の分裂の際の一瞬だけなので、この間動原体付近の接着は維持されたままである。私たちは第一分裂で染色体が両極側から引っぱられる段階で、動原体付近のRec8だけはずれないように守っている分子があるに違いないと思い、酵母でそれを探した。その結果、候補として見つかった蛋白質にシュゴシンと名づけた。減数分裂が間違いなく進むよう守っているという意味も込めた。
まず酵母でシュゴシンのはたらく場所と時期を調べてみると(図3)、分裂1回目の父由来と母由来の染色体が両極へ分かれる前に動原体近くに見られ、分かれ始めた時には消え、分裂2回目には観察できない。まさに期待通りの発現である。
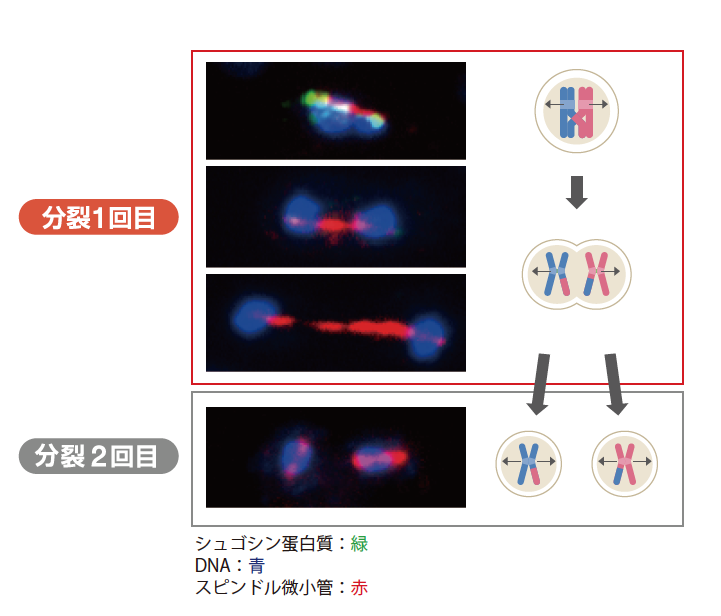
(図3) 減数分裂時のシュゴシン蛋白質の出現時期と局在
シュゴシン蛋白質は減数分裂の分裂1回目の初期にのみ現れる。染色体が細胞の両極へ向かって移動を始めると消える。
次にシュゴシンがないと減数分裂にどのような変化が起きるのかを調べた(図4)。正常な酵母の場合、減数分裂の分裂1回目の動原体付近にRec8がみられ、接着しているのに対し、シュゴシン欠損型の場合はその場所にRec8がみられず、動原体も離れていることが分かる。やはり、シュゴシンが動原体付近のRec8による接着を守っていたのである。
動原体付近の接着が維持されていることは第二分裂の際の染色体の分配にとって大切である。第二分裂前に通常はX状の染色体が離れてI状になってしまうと、細胞の両極から伸びる糸が染色体をどちらに引っぱってよいのか判断できず、しばらく停止した後にランダムに分配してしまう。つまり、シュゴシンがなく、動原体付近の接着が維持できないと、第二分裂に異常が起きるのである。
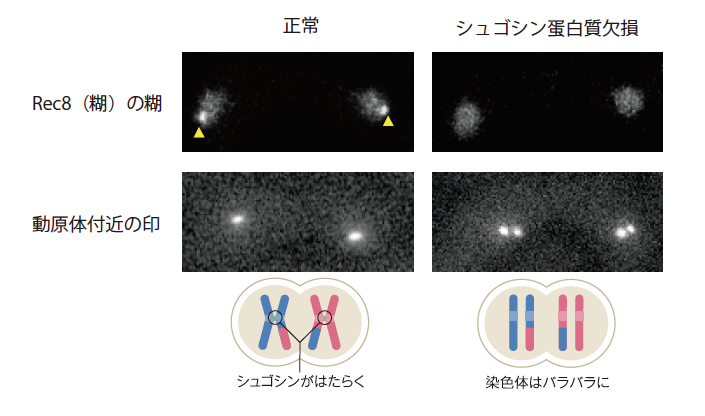
(図4) 動原体付近の接着を守るシュゴシン蛋白質
正常な減数分裂では、動原体付近の接着が守られているのに対し、シュゴシン蛋白質欠損の場合にはRec8がみられず、動原体も離れていることが分かった。
(ウ)複製した染色体を同じ方向に引っぱる
体細胞分裂では複製されX状になった染色体が反対側から引っぱられ、I状の染色体へと分かれるのに対し、減数分裂の分裂1回目では複製された父由来と母由来の染色体が組み換えによってつながり、それぞれX状のまま両極へと分かれる。複製されたX状の染色体に注目すると、体細胞分裂では反対側から引っぱられ、減数分裂では同じ方向へと引っぱられることになる。この違いは複製した染色体を接着する糊にあるのではないかと思い、減数分裂の際に染色体を接着するRec8が働かないようにした酵母を観察した(図5)。すると複製された2本の染色体が、まるで体細胞分裂の時のように、両極に分かれて1本ずつになってしまった。詳しく調べてみると、本来、体細胞分裂時に糊のはたらきをするRad21が複製した染色体の間に入り込んでいた。あるべき糊Rec8がないと、別の糊がはたらいてしまうのである。こうして、糊の蛋白質の違いが染色体分配の様式を決めていることがわかった。そこで、Rec8やRad21のはたらき方を比べたところ、体細胞分裂での糊(Rad21)は動原体の真ん中には見られず、減数分裂の分裂1回目では中央まで糊(Rec8)が埋まっていることがわかった。複製された2本の染色体が引っぱられるときに、体細胞分裂では動原体の中央が離れて両側に開き、減数分裂1回目では動原体がくっついたままと同じ方向に向くことになる。こうして動原体付近の接着の違いが染色体の運ばれる方向を決めていることが明らかになった。
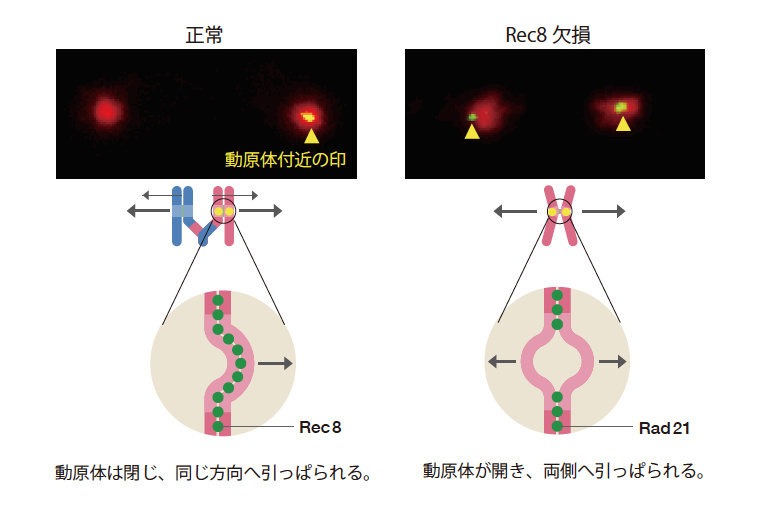
(図5) 動原体付近の形を制御するRec8
Rec8がはたらかないように酵母の細胞を操作すると、まるで体細胞分裂のように複製して接着した染色体が二つに分かれた。染色体を接着する糊も体細胞分裂型のRad21へ変わっていた。
4.奥深い減数分裂
減数分裂を特徴づける3つのしくみそれぞれについて、そこで働く分子を探し、その働き方を見てきた。この3つは、お互いに関連し合い、ひとつたりとも欠くことのできない巧妙な組み合わせで複雑な減数分裂をさせていることが分かってきた。このしくみは有性生殖をする全ての生きもので厳密保存されていることから、約10億年前にひとつの生きものに生まれそれが続いてきたと考えてよいだろう。多様性を生み出す有性生殖の重要性を示すと言える。今後は染色体の形にまで眼を向け、奥深い減数分裂のしくみ全体像を明らかにしていきたい。

渡邊嘉典(わたなべ よしのり)
1989年東京大学大学院理学系研究科博士課程修了、理学博士。日本学術振興会特別研究員、東京大学理学系研究科助手、助教授を経て現在東京大学分子生物学研究所教授。
.jpg)

.jpg)
.jpg)










