

RESEARCH
体節形成パターンを数理モデルで解く
生節足動物の体節形成は一部の例外を除き、前方部が「同時型」で後方部が「付加型」で行われていると思われていた。大学院生の金山真紀さんは、「前方部の体節が段階的な挿入で形成される」ことをクモで見出し、体節形成パターンがこれまで考えられていたよりも多様であることがわかってきた。異なる体節形成パターンを生み出す遺伝子ネットワークはどのように進化してきたのか?その問いの解明に数理モデルは有用であり、一方実験がモデルづくりに貢献する。(小田広樹)
1.数理モデルで形態進化の道筋を探る
生体内ではたらく遺伝子群はネットワークを成して発生過程を調節する。ある遺伝子のはたらきが他の遺伝子にどのように作用するのか、タンパク質間の相互作用やタンパク質とDNAの結合状態を詳しく測定する技術の発展に伴い、特定のモデル生物についてはネットワーク構造を解析できるようになった。これまでの分子生物学は、膨大な数の遺伝子の個々の機能について知見を蓄積してきた。この知見に加えて、多様な生物種それぞれが持つネットワークの機能を理解できれば、形態形成が進化してきた道筋を辿ることができるのではないだろうか。
それにしても、ゲノム上の個々の遺伝子の変異によるネットワーク構造の変化は、生きものの形態形成をどのように変容させてきたのだろうか。モデル生物以外の種で、発生過程を調節している遺伝子ネットワークの構造を実験的に解明していくことは難しい。そこで、数理モデルの計算機シミュレーションを用いる可能性を考えた。具体的には、発生過程と形態進化に関する実験的知見が豊富であり、モデルを立てやすい節足動物の体節形成に注目した。
2.節足動物に見られる多様な体節形成パターン
節足動物の胚発生では、体の前後軸方向にそって遺伝子が縞状に発現し体節形成を導くという共通の特徴がある。その中で、体節という繰り返し構造ができあがるまでの過程にはいくつかのパターンがあり、祖先種では付加型(短胚型)の発生が多く見られ、派生種では同時型(長胚型)の発生が見られる。付加型というのは、胚が伸長するとともに尾側に「1つずつ」縞が現れるものであり、同時型では胚に全ての縞がほぼ「同時に」出来上がる(図1)。ほとんどの節足動物は、付加型と同時型の発生を組み合わせた型(中間型)であると考えられている。付加型に属するヤスデなどの多足類では、同じ親から生まれた子の間でさえ体節数が異なることがあるが、同時型に属するショウジョウバエの体節数は厳密に決まっている。このように、体節数の制御の正確性にも違いがあるようだ。
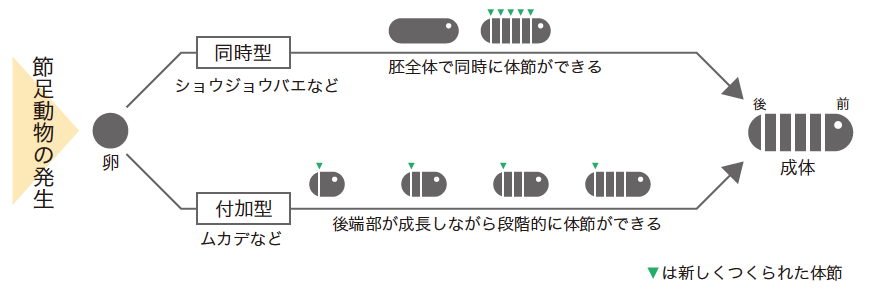
(図1) 節足動物の体節形成パターン
モデル生物であるショウジョウバエは、体節形成に関与する調節遺伝子群とそのネットワーク構造の詳細が明らかになっている。これまで調べられた限りでは、同時型の調節遺伝子群のほとんどが付加型の種と共通するので、特定の遺伝子の違いではなく遺伝子のネットワーク構造の違いが各生物の体節形成パターンを特徴づけると考えられる。
3.計算機上で進化を再現する
ネットワーク構造の変化と体節形成パターンの多様化との関係を知るには、膨大な数の節足動物の発生過程を調べ、それに関わる遺伝子ネットワークの構造を明らかにし、それぞれの体節形成パターンに特徴的なネットワーク構造を抽出していかなければならない。しかし今のところ、ネットワーク構造がわかる生物種はごく少数しかない。
そこで一旦現実から離れて、計算機を使って縞状の空間パターンを生成できるネットワークを探すことにした。生体内ではたらく転写因子が縞状の空間パターンを形成する過程を、反応拡散方程式(註1)でモデル化し、遺伝アルゴリズムと呼ばれる手法で転写因子ネットワークを計算機上で進化させたのである(図2)。つまり、計算機上で体節をもつ生きものもどきを発生させ、進化させたのである。計算機なら発生にかかる時間や世代交代が一瞬とも言える非常に短い時間で済むので、数百回の独立した進化が可能である(ただし実際には膨大な計算になるので、1日中コンピュータを稼働させる必要があるのだが)。そこで得られたたくさんのネットワークの構造と、縞形成パターンとの対応を網羅的に調べた。
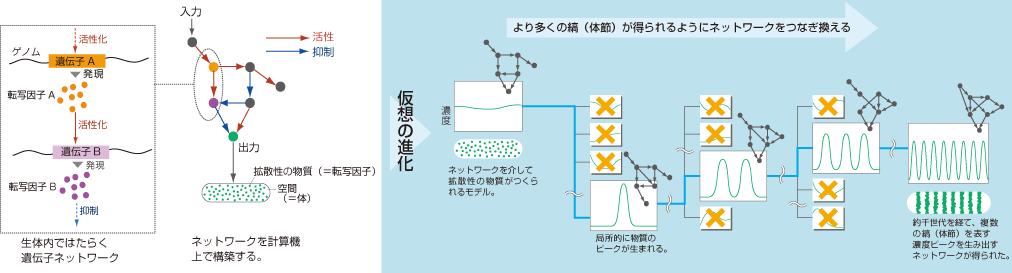
(図2)遺伝子ネットワークを計算機上で進化させる
生体内ではたらく遺伝子ネットワークをコンピュータ上で構築し(左)、ネットワークをランダムにつなぎ換え、より多くの縞が得られるように進化させる(右)。
(註1)反応拡散方程式
物質の反応と拡散を微分方程式で記述した式で、物質の非線形な反応と拡散が相互作用し、動物の皮膚の模様など多様なパターン。
4.ネットワークから見出された必須の部品DIO2
コンピュータ内での仮想の進化によって、付加型に特徴的な「一つずつ」縞を形成するネットワーク、同時型に特徴的な「同時」に縞を形成するネットワークを得ることができた。付加型のネットワークの特徴がFeed-Back Loop (FBL)という構造を必ず含むのに対し、同時型は、Feed-Forward Loop (FFL)という構造を必要とした。さらに二つを折衷した中間型は、FBLとFFL両方の部品が必要であることがわかった(図3)。FBLやFFLはバクテリアから動物にいたる転写ネットワークで多く見出されており、例えば細胞周期の進行や個体の一日周期のリズムの生成にはFBLが関わっていることが知られている。このようなFBLとFFLが、空間的リズムといえる縞の形成にどのように関わるのだろうか。
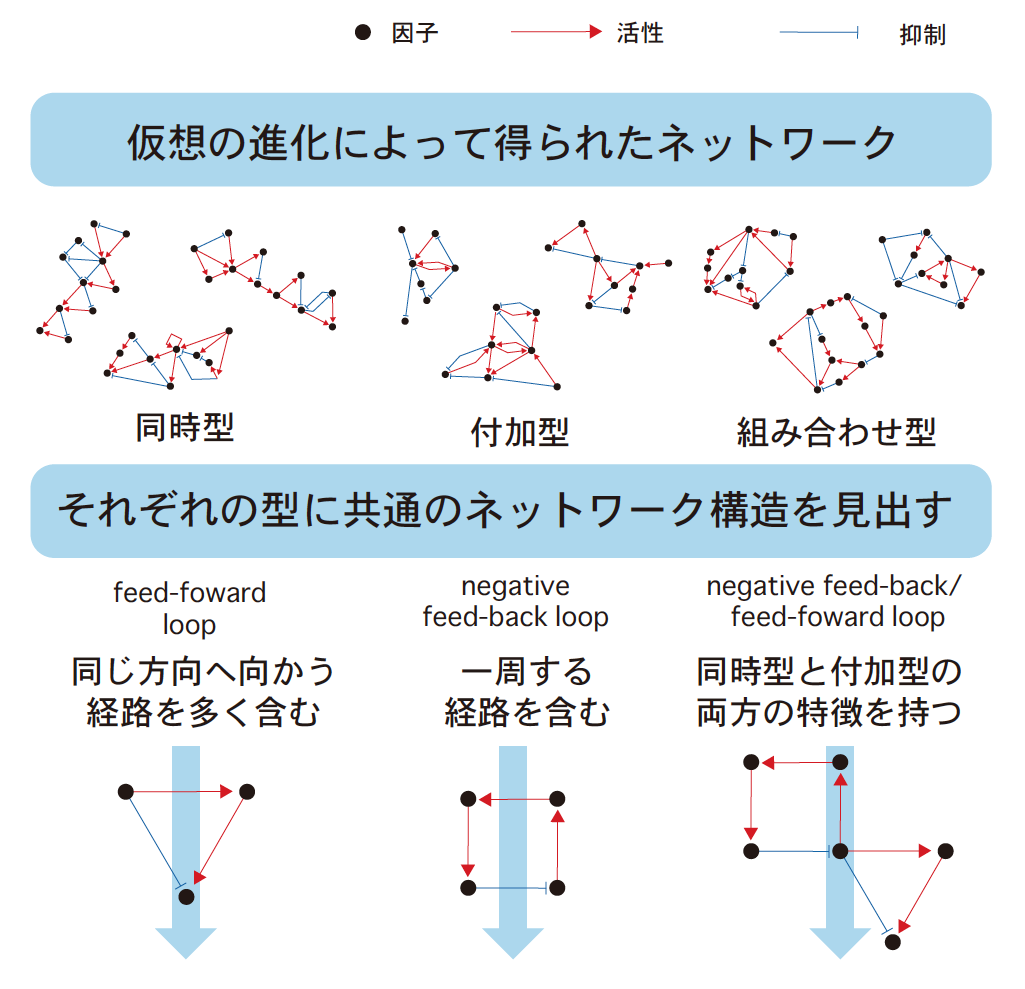
(図3) 「同時型」「付加型」「組み合わせ型」のネットワークから見出された構造
FBLが一つあれば、時間的振動を介した空間的なくり返しパターンを自発的に生成する。私たちは、このメカニズムが付加型に属する全てのネットワークで使われていることを明らかにした。しかしFBLだけでつくられる縞は、その本数の制御についての厳密さに欠け、例えば空間を倍の大きさにすると、縞の本数が倍になってしまうのだ。現実の生物では多足類がまさにそうである。
一方FFLは、この構造一つで一本の縞を生成する。すなわちFFLの数に応じて縞の数が変化するので、ネットワーク構造がストライプ本数を正確に特徴づける。したがって、空間を倍の大きさにしても、縞の本数は変わらない。FBLからなる付加型ネットワークは簡素だがいい加減であり、FFLからなる同時型ネットワークは多数の部品を組み合わせた正確な制御を行うことが見えてきたのである。
これらのネットワーク構造は、計算機上で進化させたにも関わらず、実際の生きものの遺伝子ネットワークの働きと見事に一致した。例えば、付加型ネットワークのFBLを構成する因子を潰すと時間的な振動が消えるので、まったく縞が出来なくなる。実際に付加型の甲虫トリボリウムでは、FBLの存在が示唆され、それを構成する遺伝子をはたらかなくすると縞が出来なくなる。変異体と遺伝子ネットワークを対応させる解析方法を用いると、体節形成の遺伝子ネットワークが解明されていない複数種の節足動物についても、変異遺伝子とFFLやFBLとの位置関係を推定できる。
5.仮想の進化と現実の進化をつなぐ
形態進化の道筋では、付加型の生きものから同時型の生きものが進化してきたと考えられている。そこで今は、計算機上でも付加型ネットワークから同時型ネットワークへの進化が実現するのかどうかを試み、ネットワーク構造の変化による発生過程の進化の道筋を探している。
このような研究から見て、オオヒメグモで発見された体節が分割して生じる「挿入型」は非常に興味深い。現在のモデルではこの発生様式の進化が説明できないからだ。実際の生きもののデータを用いてモデルと現実の進化の違いを検証し、モデルの質をあげていくことで、この新しい型を説明できるようにすることが次の課題である。
節足動物の体節形成は一部の例外を除き、前方部が「同時型」で後方部が「付加型」で行われていると思われていた。大学院生の金山真紀さんは、「前方部の体節が段階的な挿入で形成される」ことをクモで見出し、体節形成パターンがこれまで考えられていたよりも多様であることがわかってきた。異なる体節形成パターンを生み出す遺伝子ネットワークはどのように進化してきたのか?その問いの解明に数理モデルは有用であり、一方実験がモデルづくりに貢献する。 (小田広樹)
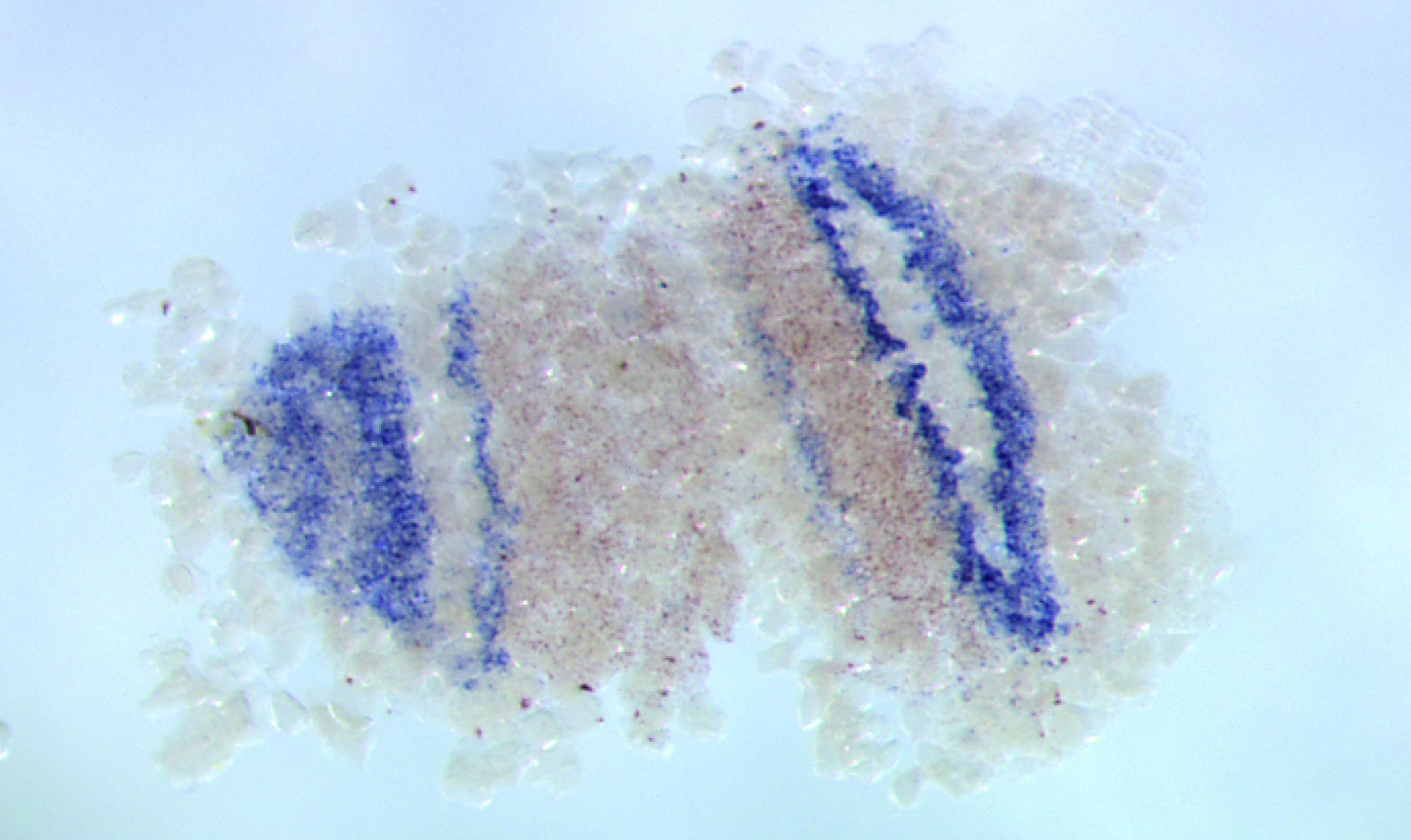
挿入によって体節が分割している。
藤本仰一(ふじもと・こういち)
2001年東京大学大学院総合文化研究科博士課程修了、学術博士。同研究科助手、科学技術振興機構ERATO複雑系生命プロジェクトグループリーダーを経て2009年より大阪大学大学院理学研究科特任准教授。
.jpg)

.jpg)
.jpg)










