

RESEARCH
トリと恐竜をつなぐ指のつくられかた
手足の形づくりに見る普遍と多様
四肢動物の指は5本が基本です。指にはそれぞれ個性があり、5本指の手の発生をみると、まず薬指にあたる第4指が腕の尺骨の先にできるので、これを基準にして第何指かを同定します。トリの手にあたる翼の先には3本の指があり、一番後の指が第4指、3本が5本指の2-3-4指に対応しているとされてきました。トリの祖先は恐竜とする証拠がいくつかあり、3本指の恐竜もいます。ところが化石でさかのぼると、恐竜は5本指から次第に後ろの指が消えて4本、3本と減ってきたようで、3本の指は1-2-3指となりトリとはつながりません。東北大学の田村宏治さんは、指を誘導する遺伝子を追い、指が発生する時間と場所を丁寧にしらべることにより、実はトリの指が恐竜と同じく1-2-3指であることを突き止めました。恐竜とトリのつながりを証明する研究です。
1.比較発生学という発想
動物は種に固有の形態をもっている。どうやってクジラやキリンやコウモリは、あんな形になるのだろう。よく見るとヒトもかなり面白い形態をしているのだが、自分のことなのであまり変わっているとは思わない。このように形を見るとき、私たちは身近な生物を基準にしてそれと違うものを「変わっている」と受け止める。ヒトの手を基準にするとコウモリの手はとても変わっているように見えるが、コウモリから見ればヒトが変わっていると見えるだろう。そこで客観的な比較をするには、共通な部分を基準にするのがよい。また、動物形態の基本は発生の過程で形成されるので、発生を追うのは、どのように共通の部分がつくられ、なにが違うと多様になるのかを理解する有効な手段になる。動物発生学の中で比較発生学と呼ばれる研究の発想は、このような単純なところから出ている。
ここでは、ニワトリの翼(前肢)と足(後肢)の先端にある指の発生の比較から、150年にわたる鳥類の恐竜起源説の論争に決着をつけた研究を通じて、四肢動物の指の発生の普遍と多様を考えてみたい。
2.発生における誘導
動物胚が発生過程で形づくりをするには、胚の中で多数の細胞が複雑な相互関係を築いていく。なかでも「誘導」と呼ばれ、ある細胞が周りの細胞に働きかけて分化させたり形を作らせたりするのは、このような相互作用の典型例である。誘導活性をもつ領域や組織を形成体(オーガナイザー)と総称する。神経を誘導する両生類の原口背唇部はシュペーマンオーガナイザーとして有名であり、他にもさまざまな例が知られている。通常、誘導現象には、形成体それ自身は誘導をかけるだけでその細胞に分化せず別の発生運命をたどるという特徴がある。
指は、種に固有であり種間で多様な形態をもつ。指の本数と個々の指の形態は指の形成体が決めるが、発生中の四肢原基(肢芽)の後端に存在するZPA(zone of polarizing activity、極性化活性帯)と呼ばれる領域がそれにあたる。ニワトリ胚を用いた移植実験で、ZPAの一部を切り出し別の肢芽の前端に移植すると指の数が倍に増える(図1)。これらの指は、ZPAの細胞から分泌されるSHHタンパク質(註1)の作用で形成され、その拡散による濃度勾配がそれぞれの指の個性をつくる。ニワトリ胚ではZPAは自分自身では指をつくらないことが実験的に示されているので、ZPAは指を誘導する形成体であるといえる。
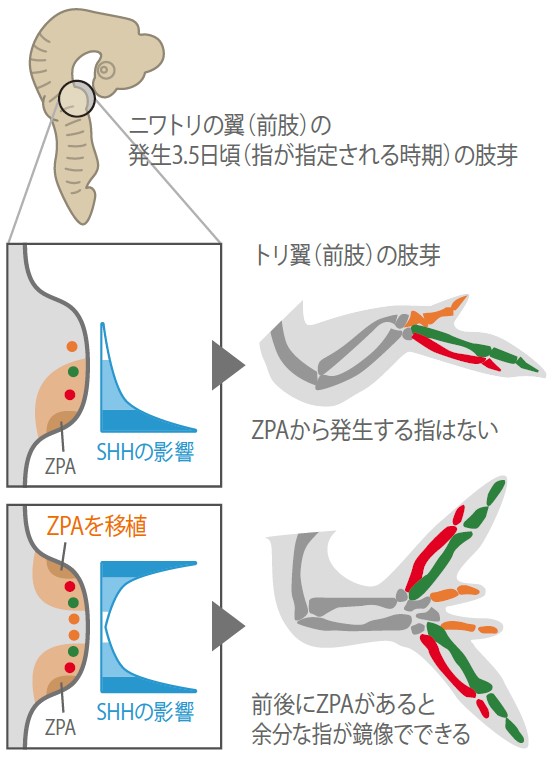
(図1)ニワトリ胚の前肢のZPAの移植実験
ニワトリ前肢では、ZPA内の細胞から作られる指はなく、ZPAをもうひとつ余分に前方に移植してみると、鏡でうつしたようにもうひと組の指ができる。
マウスでは、5本の指がそれぞれ異なる作られ方をすることが示された。まず、5本のうち最も前側の指(第1指)は、SHHの作用を全く受けずに作られる。一方、この指に続く第2指と第3指は、SHHタンパク質のシグナルを受け取った指、すなわちSHHの濃度勾配によって誘導される指である。さらに後方にある2本の指、すなわち第4指と第5指は、発生過程でshh遺伝子を発現したことのある細胞によって構成されている。実は、後方でshhを発現する領域はZPAなので、これらの2本の指はZPAからできてくることになる。
(註1) SHHタンパク質
ソニックヘッジホッグ(Sonic HedgeHog, SHH)は、発生に関わるタンパク質で、それをコードする遺伝子はshhと記載する。タンパク質が翻訳・合成されると細胞外に分泌され周辺の細胞に働きかけ、分化や形態形成をうながす。
3.ZPAの誘導作用をめぐる矛盾
私はこのマウスの研究結果に驚いた。ニワトリ胚を用いた実験では、ZPA形成体は指を誘導するだけで自身は指を作らないとされている。ところがマウスでは、後ろ側2本の指はZPAに由来するというのである。私は20年以上指づくりの共通メカニズムを研究してきたが、この点を指摘した者はいない。考えた末、この矛盾を解消するアイディアを思いついた。ニワトリの前肢には指が3本しかない。5本あるマウスの指のうち、前側の3本、すなわち第1指、第2指、第3指がニワトリの前肢の指であるとすれば、全て説明がつく。ここで私は、ニワトリ胚の後肢を用いたZPA移植実験をした。なぜなら、ニワトリの後肢は指を4本もっているので、もし私の考えが正しいなら後肢のZPAは指を作るはずだからである。
実際に、後肢のZPAを用いた移植実験では、形成される過剰指の中にZPA自身が形成する後肢の指が含まれていた(図2)。前肢のZPAは指を誘導するだけで自身は指にならないが、後肢ではZPAが指を形成することが示せたのである。そこでさらに、正常の発生で細胞を標識し、ZPA細胞の発生運命をたどったところ、前肢ではZPAの細胞は指にならないが、後肢では最も後ろ側の指はZPA細胞に由来することがわかった。マウスの指の形成を基準にすると、ニワトリの前肢の3本指は、第1指、第2指、第3指、であり、後肢の4つの指は、第1指、第2指、第3指、第4指となる。
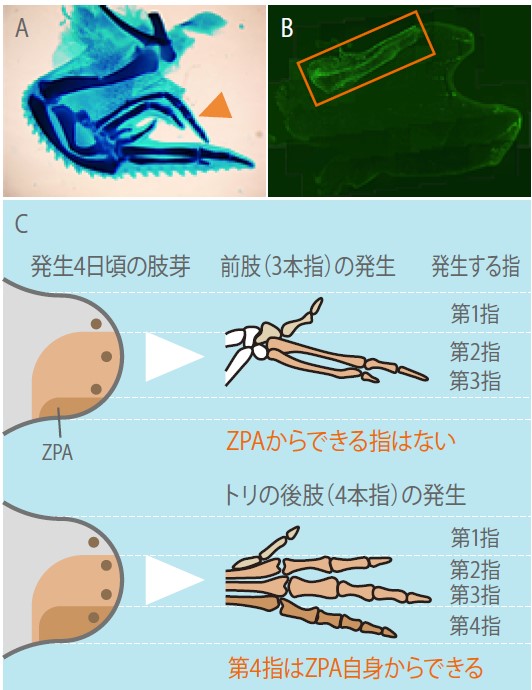
(図2)ニワトリ胚の後肢の移植実験とまとめ
A 前肢の肢芽の前方に後肢の肢芽のZPAを移植すると、図1の実験と同様に指がもうひと組できるが、一番外側の指(▲)の形状は第4指である。
B Aの実験を後肢のZPA細胞に蛍光標識して、細胞の移動のようすを見るともっとも外側の指はZPAの細胞からできていることがわかる。
C 前肢と後肢の指の形成とZPAの関係を示した。
4.2つの実験の矛盾-だから、サイエンスは面白い
私は、移植実験と細胞標識実験から鳥類の前肢の最後方の指が第3指としてZPAによって誘導されてくることを示し、鳥類の前肢の指は第1指、第2指、第3指であると主張して全ては解決したなと、ほくそ笑んでいた。
ところが、話はこれで終わらなかったのである。大学院生の野村直生が「先生の言っていることは間違っている」と言い出したからだ。彼の多点細胞標識実験(註2)によれば、前肢の3本の指の元になっている細胞は、後肢の第2指、第3指、第4指の元になっている細胞と同じ位置にあり、なおかつ最も後ろ側の指になる細胞は前肢でも後肢でもZPAの中にある、というのである。
議論と実験を重ねるうちに、実験に用いる胚の発生段階に違いがあることに気づいた。私はZPAの移植実験と細胞標識実験を、ニワトリ胚の発生段階22(3日半)という、ちょうど指形成が開始する時期を選んで行っていた。ところが野村は、指が作られ始める前で、前腕部を形成中の発生段階のより早い胚、発生段階20(3日)の胚を実験に用いていたのである。 この発生段階20から22まではおよそ16時間程度。もし両者の実験データが正しいとしたら、この短い時間の中でずれが生じているはずだ。
実験の結果は予想通り、後肢では16時間経ってもZPAの中に留まっていた指になる細胞群が、前肢ではZPAの外に出てしまっていたのである(図3)。結果として指が形成される頃には、前肢の最も後ろ側の指を形成する細胞はZPAの外側でSHHの誘導を受け、第3指を形成する。
こうして全ての矛盾が解けた。そして私たちは「発生学的にみて、鳥類の前肢の指は第1指、第2指、第3指である。ただしこれは指が3本しかないために発生のごく初期にずれ(フレームシフト)が起きてしまうことが原因となっている。」と主張することになった。

(図3) ZPAの細胞(SHHを発現)の位置と指の関係
前肢では、発生段階20(発生3日目)と発生段階22(発生3.3日目)の間でSHHの発現が変化し、ZPAではなくなった細胞が誘導されて第3指をつくる。後肢では、ZPAの細胞はそのまま第4指をつくる。
(註2) 多点細胞標識実験
発生中の異なる細胞群を複数の蛍光色素などで標識し、細胞の移動や形成する組織などを追跡する実験方法。
5.ニワトリの指と恐竜の指
実は私は、鳥類の前肢の指の番号について15年近く考え続けていた。これが鳥類恐竜起源説を支持しない大問題となる「鳥類の前肢の指問題」だからである。
鳥類と爬虫類の近縁関係は強く、鳥類が爬虫類の一部から派生したらしいことは、比較形態学でも古生物学でもゲノムサイエンスでも明らかだ。なかでも獣脚類恐竜という肉食恐竜の一部が最も鳥類に近縁とされ、羽毛獣脚類恐竜の発見などから、古生物学的には鳥類恐竜起源説はほぼ確定したとされている。指に関してみてみると、古い時代の恐竜化石の前肢には5本の指があるので、これを基準に獣脚類恐竜の前肢骨格を化石年代順に並べてみると、5本だった指のうち後ろ側の2本が短く痕跡的となり、前側の3本が残っていく様子が見てとれる(図4)。したがって古生物学的証拠からは、鳥類の前肢の指は残りの第1指、第2指、第3指である、となる。
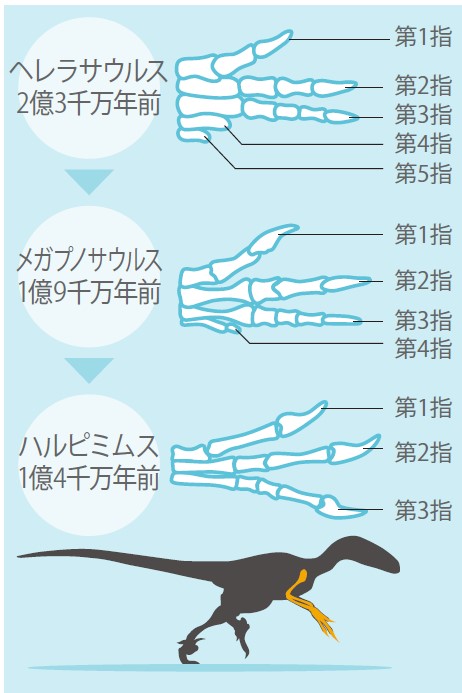
(図4)恐竜の化石からみる指の数の変化
時代を経るにしたがって、指が後ろから順に減っている。
ところが発生学者たちは長らく鳥類の前肢の指の番号を第2指、第3指、第4指としており、発生学のあらゆる教科書には第2指、第3指、第4指と書かれている(図5)。発生学者の中にはこの矛盾を理由の一つに、鳥類と恐竜の類似は収斂であり、鳥類は恐竜以前に主竜類から分枝した別の主竜目爬虫類に起源するという主張さえあった。こうして、発生学者は鳥類恐竜起源説を受け入れていないという空気がつくられてきた。
今回私たちが示した、鳥類の前肢の指は発生学的に見ても第1指、第2指、第3指であるという答えは、このパラドクスを解消する結果となった。始祖鳥や3本指の獣脚類の発生過程を見ることはできないが、それらはニワトリがそうであるように、前肢の指を発生するときに初期フレームシフトを起こして、第3指がZPAの外に形成されているはずである。つまり、ニワトリとマウスの発生の比較が、鳥類と共通した恐竜の形態の理解にもつながったのである。これこそ、比較発生学的発想が目指すひとつのパラダイムだ。
大きな問題を解決したことは嬉しいが、実は私にとって最も重要なのは、ZPAは形成体か否かという問題に対して、それが第2指と第3指の形成体であるという解答を出すことができたことだ。ニワトリの前肢という実はとても特殊な形態を見ていたために異なって見えた現象も、この四肢動物に普遍的な方法で説明できる。ZPAの発見から40年以上、まだこんなに分かっていないことがある。まだまだやれる、あらためてそんなことを思う。
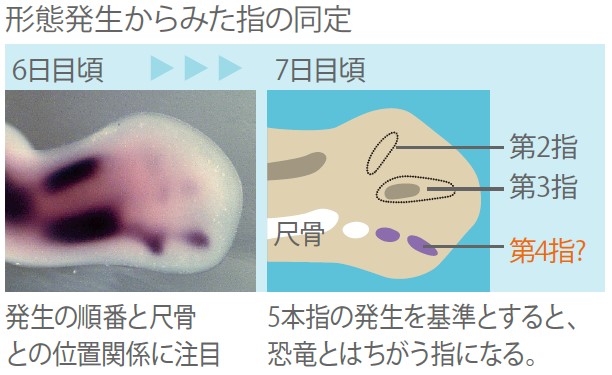
(図5) 従来発生学で考えられて来た指形成の順序
5本指の生物では、第4指が一番始めに尺骨の前にできるため、それにならって尺骨の前に形成される指を第4指とすると、その前にできる指は第2、3指になる。この考え方は発生学では広く浸透し、教科書などにも記載されてきた。
田村宏治(たむら・こうじ)
1993年東北大学理学部大学院博士課程修了。博士(理学)。東北大学大学院理学研究科助手、米国ソーク研究所ポスドク、1999年より東北大学大学院理学研究科助教授、生命科学研究科助教授を経て、2007年より同大学院教授。
.jpg)

.jpg)
.jpg)










