

PERSPECTIVE
ゲノムから見る生命誌の時間
生命誌は、生命誕生から現在そして未来にむけて続く物語です。ゲノムをもつ細胞である最初の生きものから、あらゆる生きものが生まれました。暮らしの場や周りの生きものとの関わりの中で、もっているゲノムを使い、変化を受け入れ、続いてきた子孫がわたしたちです。ゲノムにはその物語が綴られています。生きもの進化の道のりをゲノムから見ましょう。
1.生きものの誕生
生きものの始まりの時を私たちは見ることはできませんが、研究や発見を通して実態に迫ってきました。生きものの基本は、代謝し、複製し、分裂することです。代謝の起源は、原始の堆積岩の中の私たちの体をつくる有機物の痕跡によって知ることができます。およそ38億年前のイスア深海に由来する岩石に生きものがつくったと考えられる化合物が含まれているので、生命誕生はそれ以前に遡ることができます。
最初の生きものは、バクテリア(真正細菌)やアーキア(古細菌)などの原核生物と考えられています。私たちヒトを含む、目に見える生きものである真核生物は、バクテリアとアーキアとの共生から生まれたことが、多くの証拠から言われているからです。生きものの進化は、バクテリアとアーキアの共通祖先の生きものLUCA(Last Universal Common Ancestor:最後の普遍的な共通祖先) から始まることになります。40億年前ともされる古い出来事であり、それ以外にもまだ知られていない生きものがいたかもしれません。LUCAがどのような生きものだったのかは、難しい問いです。
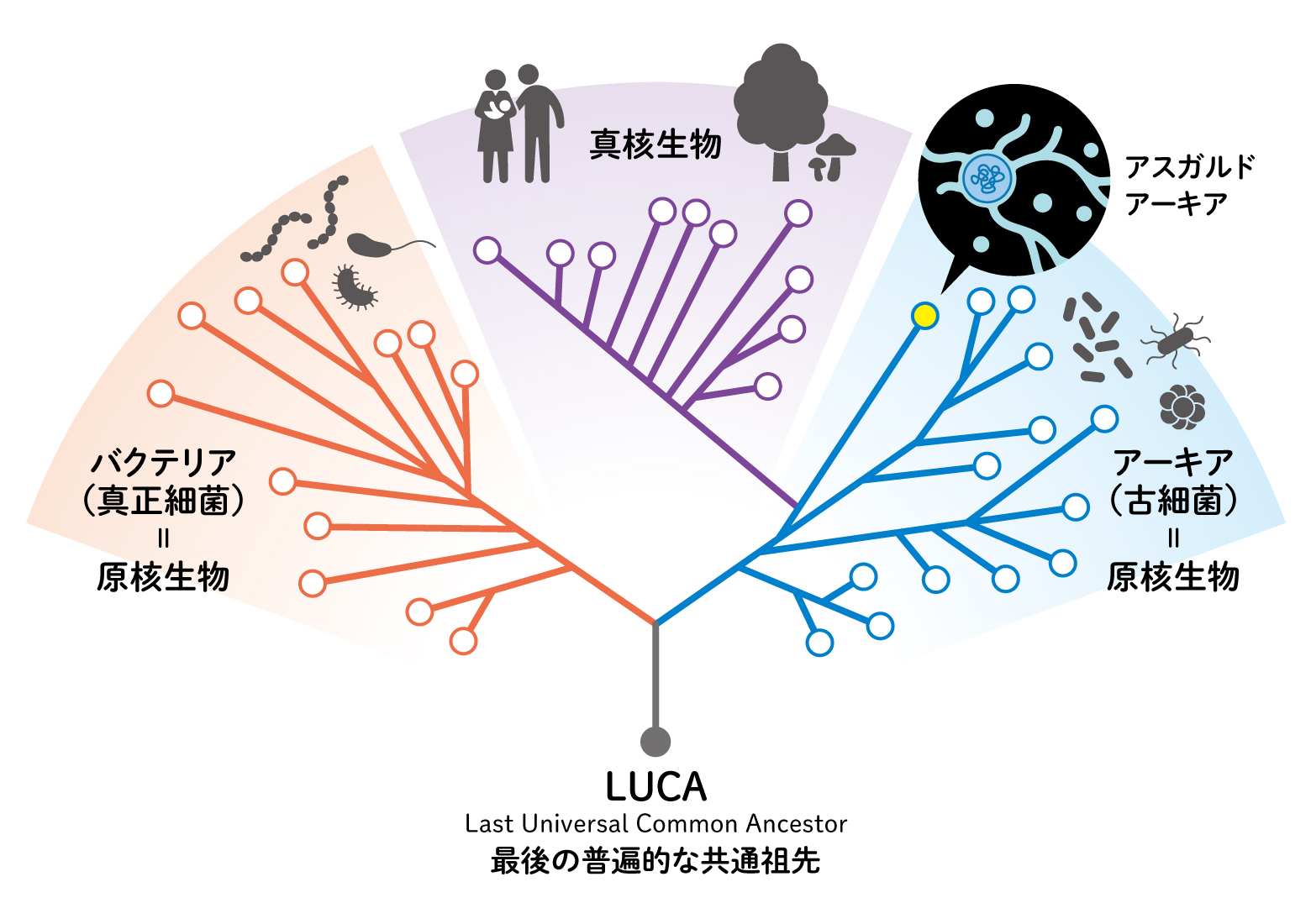
(図1) 3つのドメイン
現在の生きものは、バクテリア、アーキアの原核生物と真核生物の3つのドメインに分けられます。
2.真核生物の誕生
真核細胞は、アーキアがバクテリアを体内で飼い慣らすことでできた、という考えは「細胞内共生説」として広く知られています。真核細胞がもつ細胞小器官であるミトコンドリアは、α-プロテオバクテリア、葉緑体は、シアノバクテリアの共生です。ミトコンドリアと葉緑体が、それぞれDNAをもっていること、飲み込まれて膜で包まれたように二重の膜をもつことが、その根拠とされました。最近では、高温高圧などの過酷な環境に生きるアーキアが、バクテリアを飲み込めるほど大きな口をもっていないこと、真核生物の外側の細胞膜はバクテリアに近いことなどから、アーキアは飲み込んだのではなくバクテリアと融合したという考えがでてきています。
2015年、熱水が噴き出す深海から新種のアーキアが発見され、真核生物の祖先として注目を集めました。アスガルドアーキアと名付けられたそのアーキアの仲間は、真核生物特有の細胞骨格などの遺伝子をもっており、触手を伸ばしたり、小胞を出したり、周辺のバクテリアと物質をやりとりする様子が観察されました。そこで、アーキアは、毒である酸素を取り除いてくれるので、バクテリアと共生したのではなく、酸素のない深海で水素や栄養などを得るために共生したという考えに変わってきています。共生のおかげで酸素がある場所でも生きることができたので、真核生物につながったという考えです。
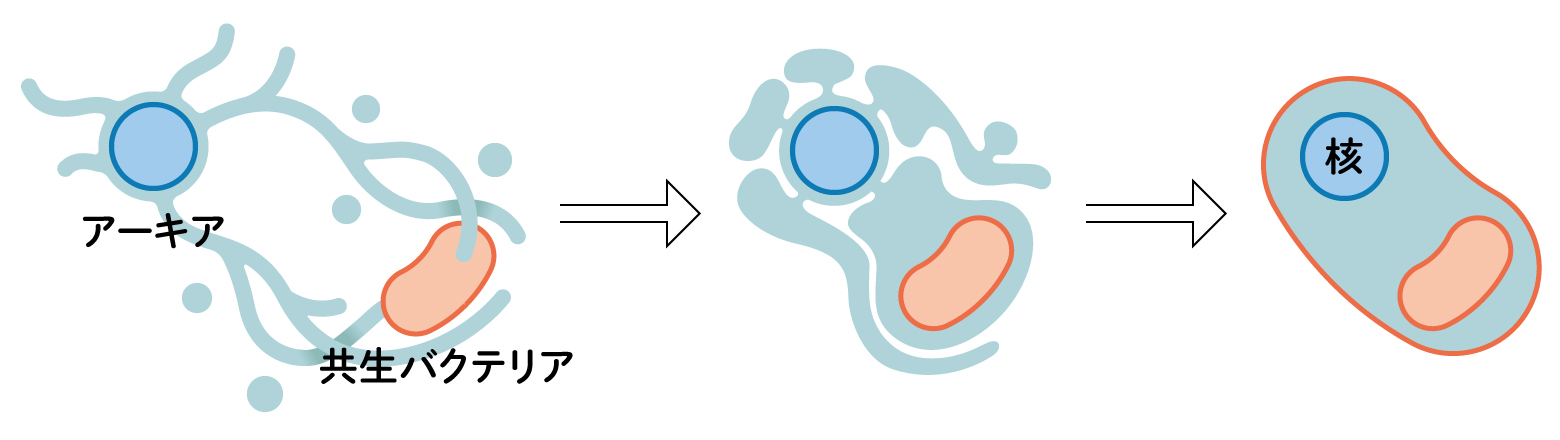
(図2) 真核生物の誕生
アーキアと共生バクテリアは、栄養やエネルギー源の水素などを交換し、次第に一体化したようです。共生したバクテリアがエネルギーを生産することで、大きく複雑な細胞になれました。核の由来はわかっていませんが、バクテリアの遺伝子のほとんどは核に移動しました。
原核生物のゲノムは、DNAが輪になっており、遺伝子がほぼ隙間なく並んでいます。他の生きものと遺伝子をやりとりすることが得意で、環境が変化したときに役立つ遺伝子をもっていたものが生き残ります。
真核生物は、エネルギーをつくるミトコンドリアを手に入れて細胞を大きくし、ゲノムを倍に増やしました。ゲノムの形も両端のあるヒモ(染色体)に変わり、遺伝子と遺伝子の間だけではなく、遺伝子の途中にもイントロンと呼ばれる隙間の配列が入っています。この真核生物のゲノムは、外から遺伝子を入れるよりも、同じもの同士でゲノムを組み換えて多様な子孫を作り、変化に対応しました。これが、有性生殖の起源とされますが、性の意味にはさまざまな説がありまだまだ研究途上です。 [1]
3.多細胞動物の起源
真核生物の多くは今も単細胞で、ほとんどが水中に暮らす微生物です。多細胞になった生きものは、動物、植物、菌類。アメーバの仲間の粘菌は生活環の中で多細胞になる時があります。カビやキノコなどの菌類は、植物に近いように思われていましたが、ゲノムを比較したところ、実は動物に近いことがわかり、オピストコンタという同じグループに分類されています。つまり、菌類と動物は共通の祖先から分かれてそれぞれに進化したということです。現存の単細胞生物のゲノムを比較して、動物と菌類の祖先を推測したところ、動物の祖先に近い生きものは、多細胞になる前から、多細胞動物の特徴を示す遺伝子をもっていたようです。また、遺伝子の数を増やし、遺伝子を重複させたり、つなげたりして新しい遺伝子をつくる傾向が見られました。大きく複雑な体をつくる準備と考えられます。一方で、キノコの祖先に近い仲間では、さまざまな代謝に関わる遺伝子を増やし、他の生物から取り込んだものもありました。動物ほど単細胞からの跳躍はありませんが、キノコは分解者として生態系の循環に関わり、また、毒などさまざまな化合物を合成することにも長けています。多細胞になる前から、動物には細胞同士が協力してはたらくさまざまな能力、キノコにはキノコらしさを発揮する能力を、それぞれ準備することで進化につながったようです。[2]
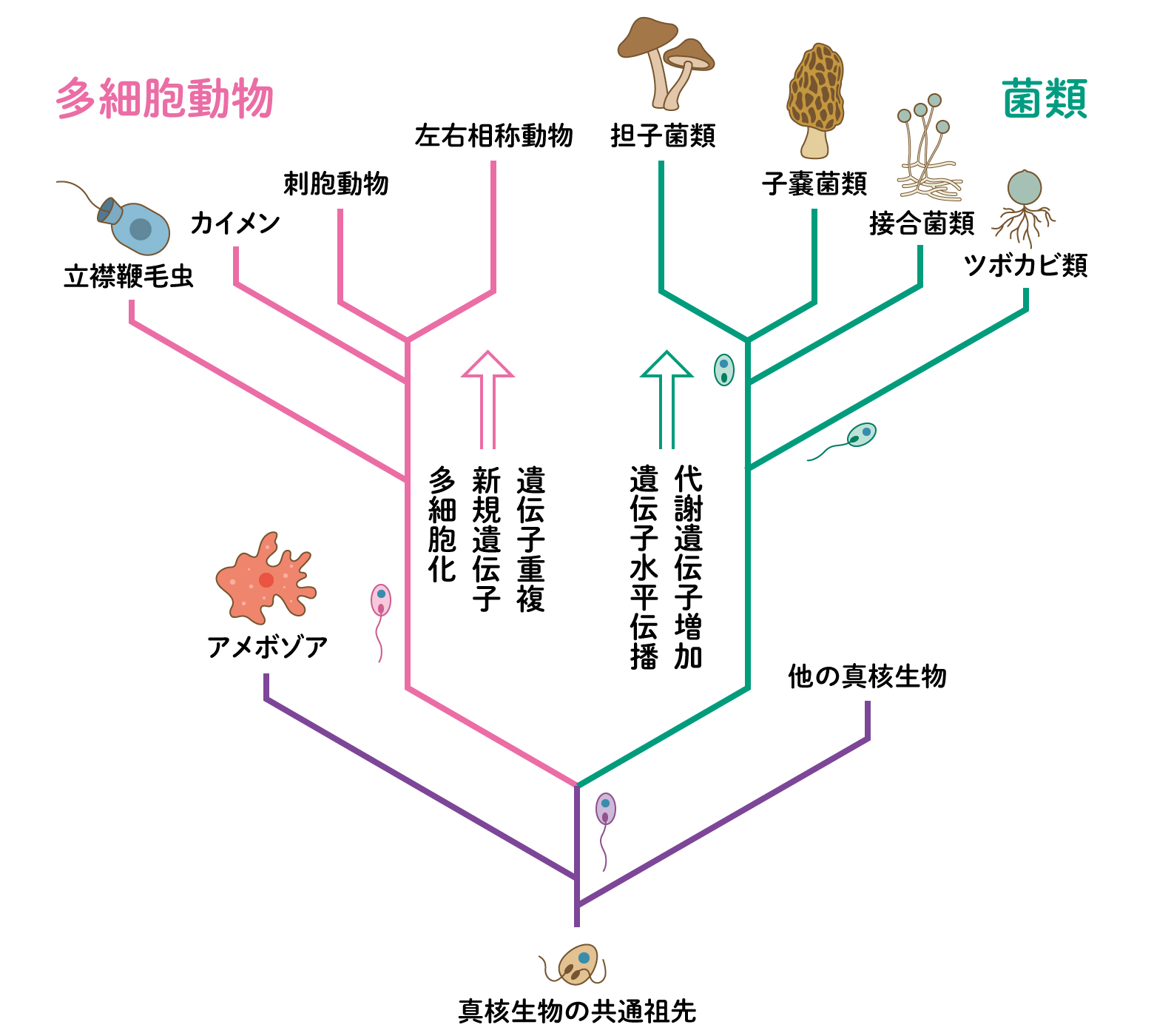
(図3) 動物と菌類の進化
動物と菌類の共通祖先から、動物に向かう生きもの菌類に向かう生きものに分かれました。
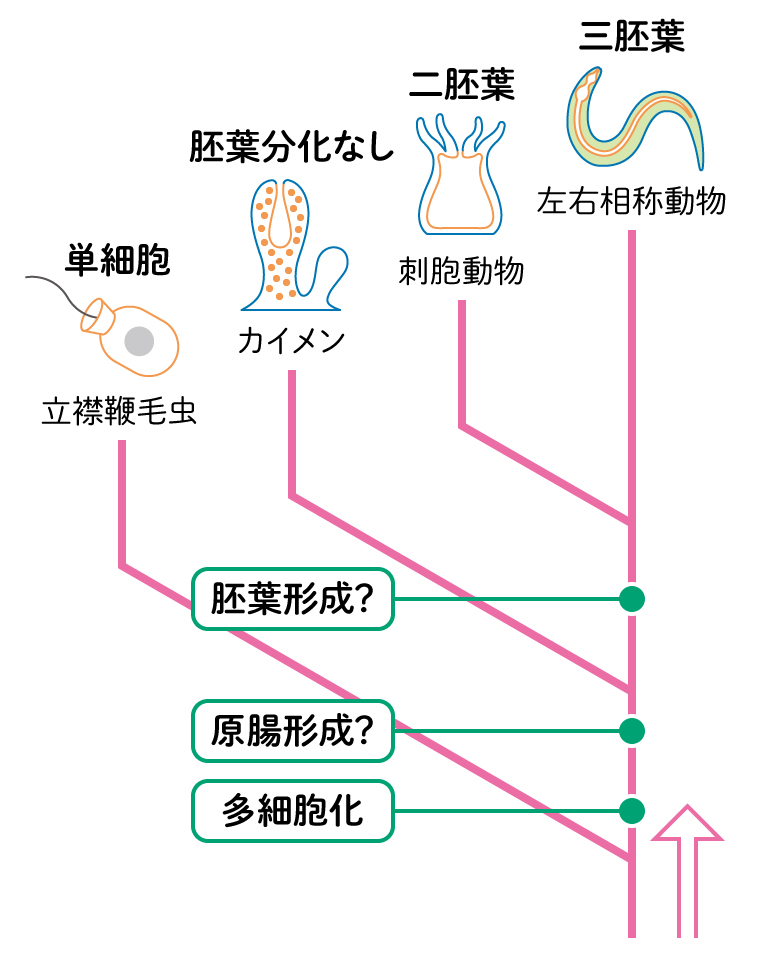
多細胞動物の発生
多細胞動物は、卵から発生で体をつくります。発生が始まると、外側に表皮や神経になる外胚葉、内側に消化管になる内胚葉、その間に中胚葉と3種の細胞に分かれます。最も原始的とされる海綿動物は、細胞が移動して位置関係がなくなり胚葉はできません。次に単純とされるクラゲやサンゴの仲間の刺胞動物は、外胚葉と内胚葉の2胚葉に分かれますが、中胚葉に似た性質ももつようです。6億年ほど前の丸い卵のような化石に胚葉らしい構造が見えていますが、どんな生きものかはわかっていません。[3]
4.多細胞動物の躍進
多細胞動物は5億4千万年前のカンブリア紀に、突然、目に見える大きさに進化しました。当時の地層にさまざまな海洋動物が見つかり「カンブリア爆発」と呼ばれます。しかし、化石が見つかるということはその頃すでにいた証拠ですから、いつどこで生まれたのかはわかりません。およそ3千万年を遡ったエディアカラ紀にみつかる化石では、生きものたちはずっと単純な姿をしていました。カンブリアの生きものは、モンスターと呼ばれるように形は奇妙ですが、今の生きものにどこか似て、目があり、口があり、足があるものもいます。
ゲノムの解析技術が進み、全真核生物のゲノムを解読するプロジェクトが行われています。そこで、これまでにわかった主な多細胞動物のゲノムを比較して、似ているところを並べると、多細胞動物の共通祖先のゲノムが復元できました。共通祖先のゲノムをピースに分けて、組み換えたり、重複したりすると、今の動物のゲノムができるのです。それぞれのピースが目や口や足、そして今いる動物に共通するしくみを作るしかけに関わっているかもしれません。カンブリア爆発は、ある時、共通祖先のゲノムをシャッフルするような事件が起きて、さまざまな形が現れたと考えることができそうな研究がでてきています。[4]
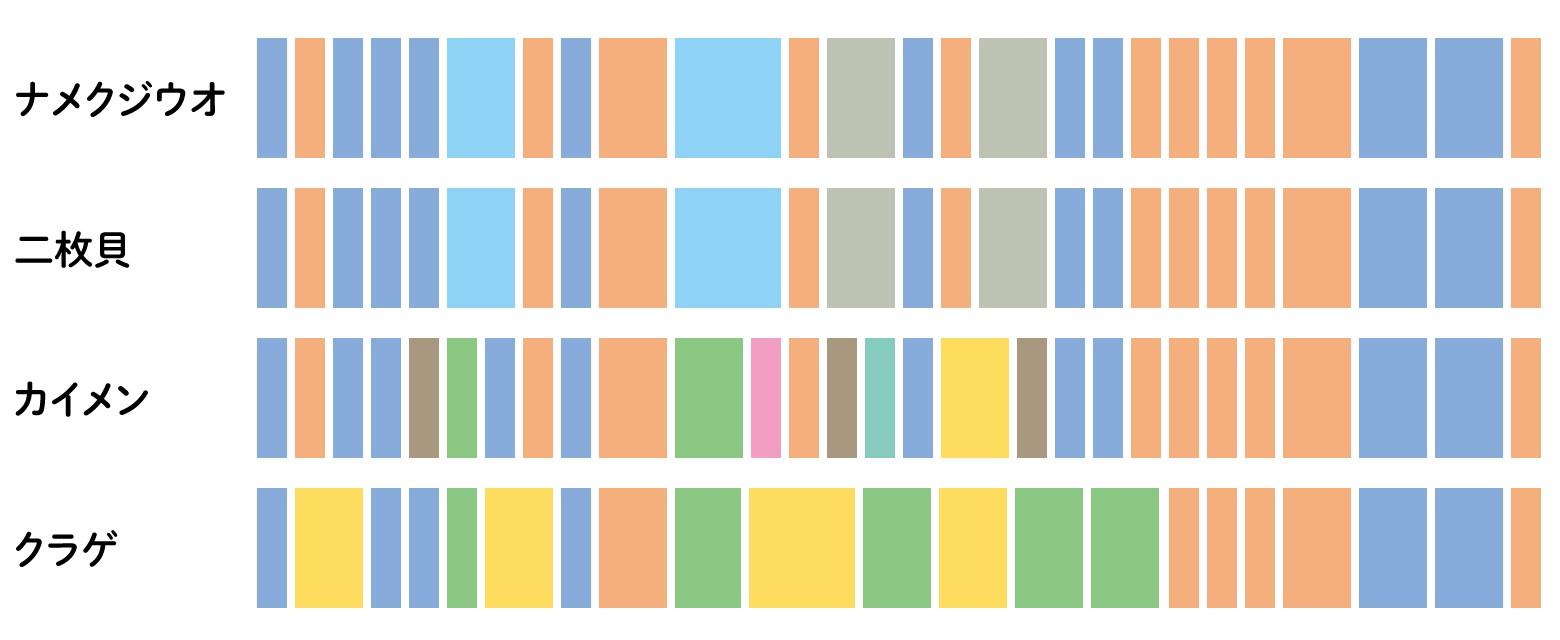
(図4) 多細胞動物で保存されているゲノムブロックの比較
動物の間でゲノムを比較して、よく似たブロックを並べました。カイメンを中心に二胚葉(クラゲ)、三胚葉(二枚貝、ナメクジウオ)と変化しており、それぞれの進化との関わりを想像させます。
文献[4]のFigure3より改変。
環境と進化
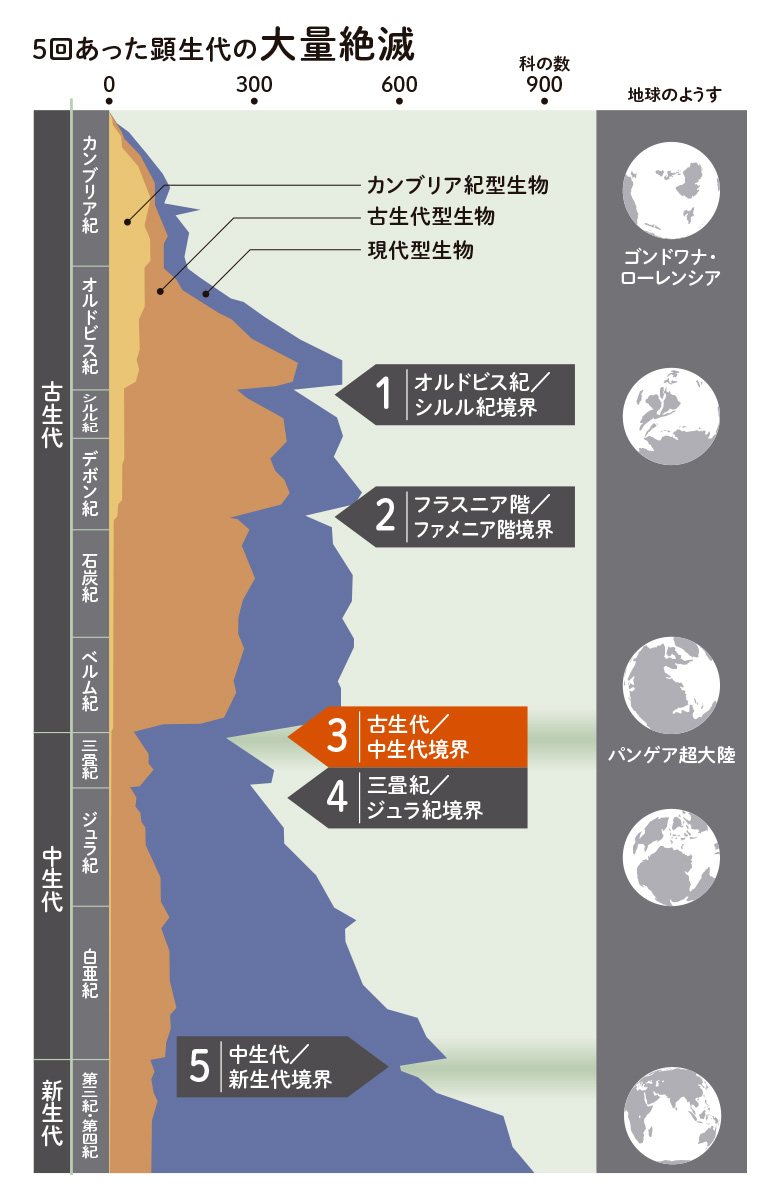
カンブリア爆発で、現在の動物のほぼ全ての門*に分類される生きものが登場し、今も続いています。つまり、ゲノムからつくられる動物の体の基本は準備できたといえるでしょう。現在までさまざまな生きものが生まれ、消えて、今の生物の多様性が築かれました。そこには、ゲノムに記されたしくみだけではなく、生きる世界、環境が大きく関わります。地球の環境は、太陽や飛来する天体など外からの影響や地球内部の変動によって劇的に変化することがわかってきました。温度の変化、酸素や二酸化炭素などの組成、陸地と海の関係や火山活動、気象状態など、生きものが生きることは、地球環境の変化に耐えることです。過去の大きな変動は、生きものに大きな打撃を与え、5回の大絶滅を引き起こしましたが、生き残った生きものから、新しい形質が出現して、ここまで続いてきたのです。
*脊椎動物門、節足動物門など33門
5.脊椎動物のゲノム
カンブリア時代の脊椎動物の祖先は、現在のナメクジウオのような姿でした。シルル紀の終わり頃まではアゴもヒレもない大人しい生きもので、ヤツメウナギなどの円口類が今では残るだけです。やがて、ウミサソリなどの無脊椎動物が繁栄する海で、ヒレで泳ぎ、アゴをもつすばしこい魚類が現れ、続くデボン紀には一気に多様化し、魚の時代を迎えました。
ナメクジウオと脊椎動物のゲノムを比較すると、脊椎動物ではナメクジウオの4倍に増えていました。ナメクジウオが1つもっている遺伝子が4つに増えていたのです。ヤツメウナギのゲノムを調べると、ナメクジウオから8倍に増えていました。そこで他の脊椎動物のゲノムとも比較すると、ヤツメウナギとアゴをもつ魚の共通祖先でまず2倍、ヤツメウナギと分かれた後の脊椎動物でもう2倍、魚の仲間でさらに2倍、ヤツメウナギだけで4倍増えたことがわかりました。[5]
ゲノムが倍化すると、遺伝子が増え、増えた遺伝子が新しい機能をもつことで、新しい形や機能がつくられると考えられます。古代魚といわれるポリプテルスの仲間は、足の関節を動かす遺伝子をヒレで使い、肺機能の遺伝子や、空気中の分子を嗅ぎ分ける嗅覚の遺伝子をもっています。おそらく現在の大多数の魚である真骨魚類と古代魚との共通祖先は、肺や肉鰭をもっており、真骨魚類が失ったのでしょう。肺は真骨魚類では浮き袋になったとされます。進化の過程では失うことも適応であり、ヤツメウナギのようにあまり変わらずに生きていくことも、もてるものを使って新しい環境に挑戦することも、さまざまな方法があるのです。[6]
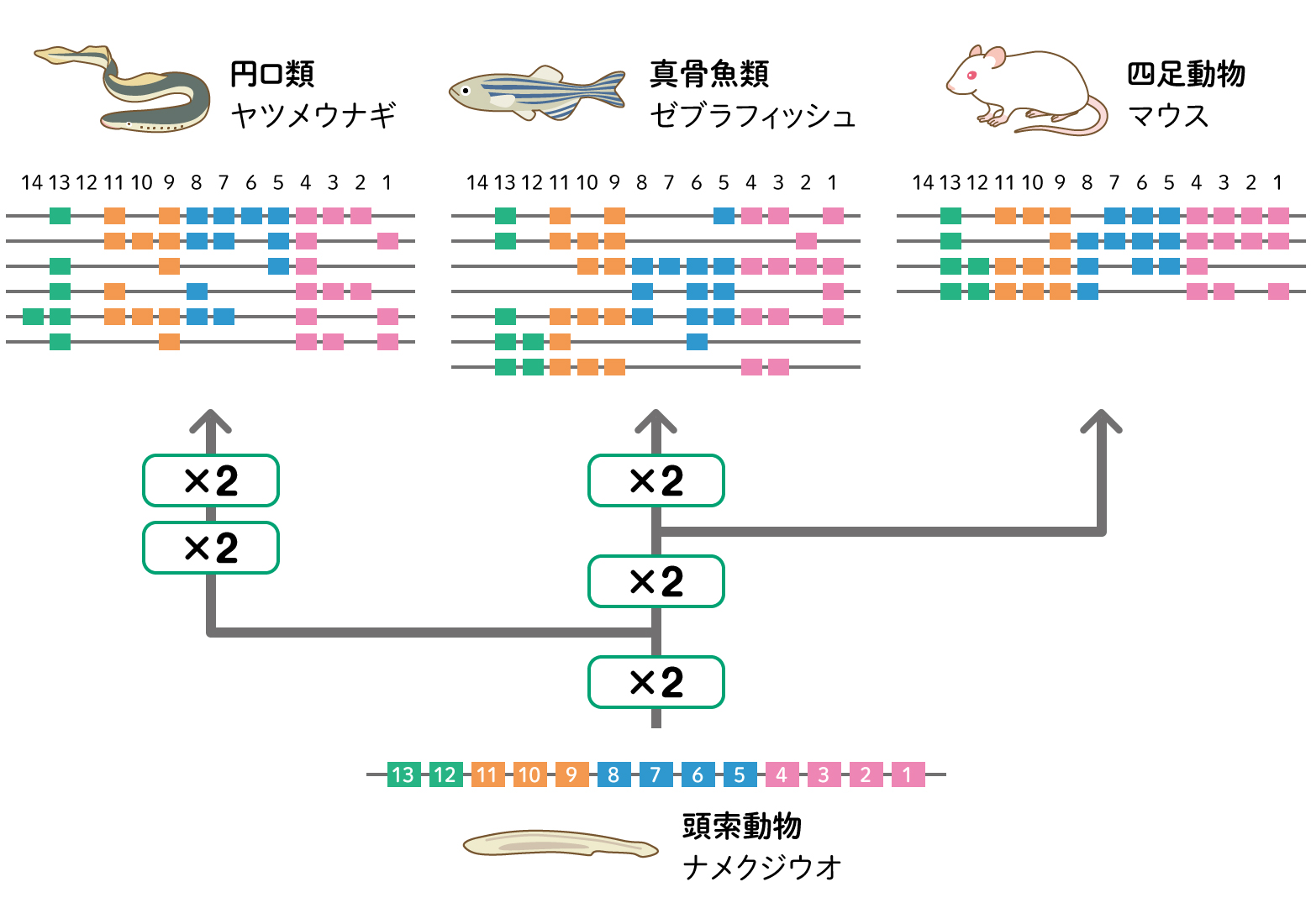
(図5) HOX遺伝子の倍化
HOX遺伝子クラスターは、ナメクジウオが1セットもっており、ゲノム倍化を調べる指標としてつかわれます。頭から尻尾のからだの位置の番地を示しますが、手足の指や心臓など部位や器官の位置や形にもつかわれています。
ゲノムの倍化は、植物にも見られ、裸子植物と被子植物の共通祖先、被子植物でそれぞれ2倍になり、その後多くの系統でさらに倍化しています。

上陸の物語
脊椎動物が陸上に現れたのは、デボン期終盤、海洋生物の80パーセントが絶滅した2度目頃(3億7千万年前)です。その頃、巨大化した植物が森林となり、枯死した後は陸地を覆い、水辺にも堆積したようです。水中ではバクテリアが有機物を分解するのに酸素を使うので、魚には息苦しい場所になりました。そこで、水際に上がり、やがて陸で暮らすことを選んだかもしれないのです。陸上で体を支えることになる四肢は、朽ちた植物をかき分け、頭を上げて呼吸するのに役立っていたという考えもあります。上陸を果たした脊椎動物は両生類とされますが、現在のカエルやイモリなどとの関係はわかっていません。
爬虫類と鳥類、哺乳類の祖先は、水辺を離れて暮らすため、卵を陸で孵すしくみとして胚をつつむ膜をもつ羊膜卵を産むようになったので、羊膜類と呼ばれます。羊膜類は、頭の骨の穴の数から、哺乳類の祖先が単弓類、爬虫類の祖先は双弓類と呼ばれました。
地球史上、最大の絶滅と言われるペルム紀―三畳紀の絶滅期には、陸上も海中も90パーセント以上の生きものが絶滅したとされています。この絶滅で、三畳紀の陸上で多様化し、生態系の頂点に立っていた単弓類のほとんどがいなくなりました。その頃の単弓類は、爬虫類型哺乳類とも呼ばれ、今の哺乳類のように体を包む毛や飛び出した耳や鼻をもっていませんでした。ここでわずかに生き残った種がつないだ中生代は、爬虫類の仲間から恐竜が現れて、主役が交代したと考えられています。

淡水魚類が陸上に進出し、両生類、さらにトカゲ型の初期の羊膜類が現れました。その後、ペルム紀末の大絶滅が起こり、単弓類のリストロサウルスが生き延びた様子が描かれています。(「生きもの上陸大作戦」展示の上陸絵巻より)
6.羊膜類のゲノム
現在の爬虫類と鳥類のゲノムを比較して恐竜のゲノムを推測すると、両者で共通する大きな染色体と小さな多数の染色体に分かれていたことが考えられました。染色体が組み換わりやすいこの構造が、およそ1億5千万年近く白亜紀の末まで、恐竜が多様な種を生み出し、大型の恐竜が絶滅した後に鳥として繁栄したという考えがあります。哺乳類では、卵を産む原始的な単孔類の染色体は爬虫類や鳥に似ていますが、子供を袋で育てる有袋類では大きくつながった少数の染色体をもっています。哺乳類では霊長目、食肉目など種類によって組み合わせが異なります。もっている遺伝子はほとんど同じなので、染色体の違いが種を分けているのでしょう。[7]
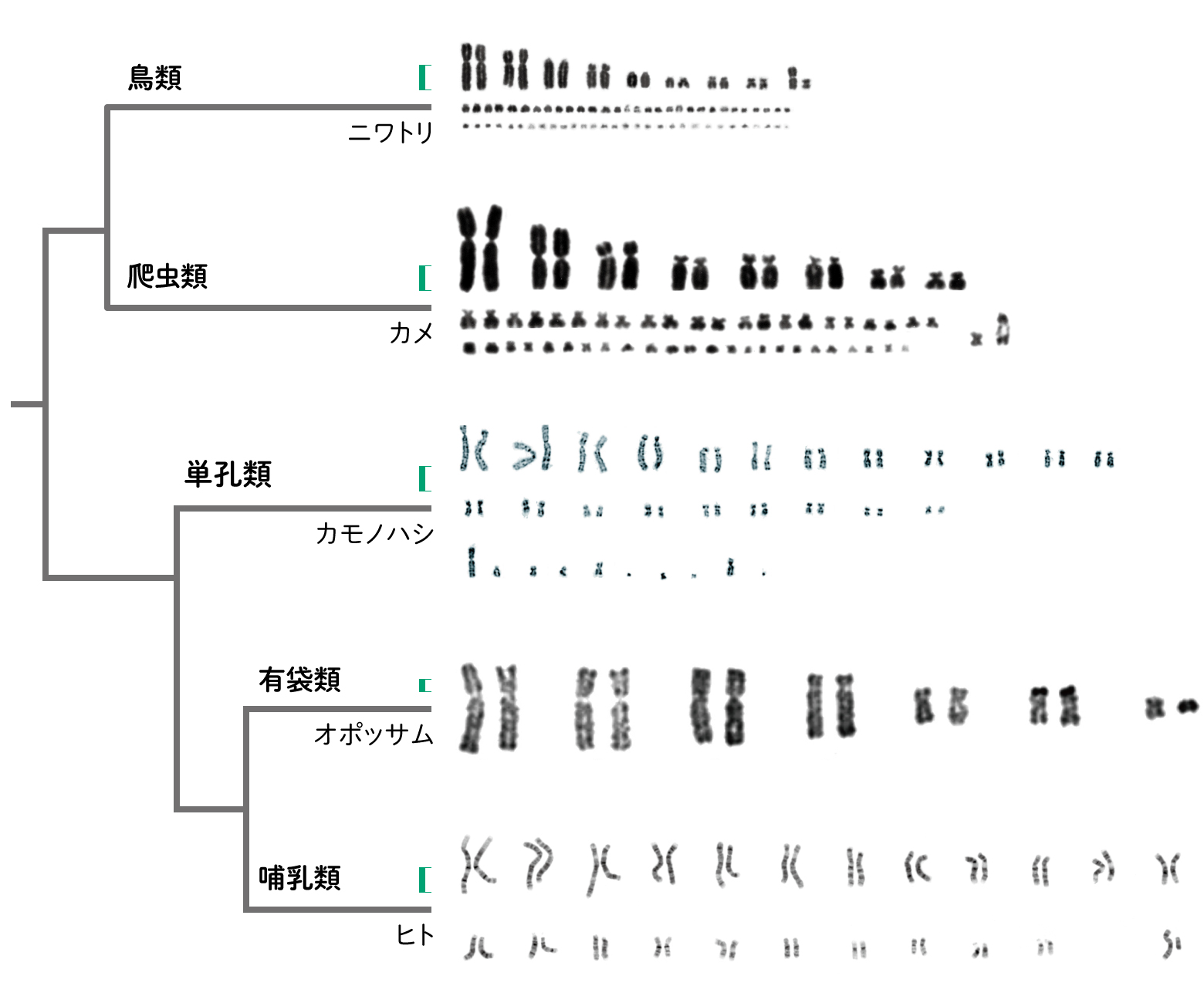
(図6) 染色体構造の比較
爬虫類、鳥、単孔類は大きな染色体(マクロ染色体)と小さな染色体(ミクロ染色体)をもちますが、獣類(有袋類、有胎盤類)では、マクロ染色体のみになります。
スケールバー(緑)を示しましたが(1億塩基程度)、染色体は凝集しているので縮尺の違いの参考としてください。
ニワトリ、カメ(スッポン)文献[8]のFigure_S1、カモノハシ 文献[9]のFigure 6、オポッサム 文献[10]のFigeure1、ヒトNational Human Genome Research Institute より改変。
恐竜の繁栄
恐竜の繁栄の原因としてしばしば挙げられるのが、気嚢による呼吸の発達です。大気中の酸素が減少した絶滅事変の後、効率的に酸素を取り込むことができ、しかも骨にも空気の道をつくることで骨を軽くすることができました。最大40メートルの巨体をもつ竜脚類は、空気を貯めた軽い骨でできた長い首で、呼吸も不自由なく首を動かして大量の植物を食べていたと考えられています。この巨大化を可能にしたしくみが、後に軽い体で飛ぶことを可能にした鳥の進化につながるのですから、何が将来役に立つのかという考えは、生きもの進化には通じません。一方、大型の恐竜が活動する昼間を避けて、哺乳類は夜行性になったと考えられていますが、三畳紀に生き残っていたものは土の中で暮らしていたとも言われ、暗いところへの適応がつながったのかもしれません。
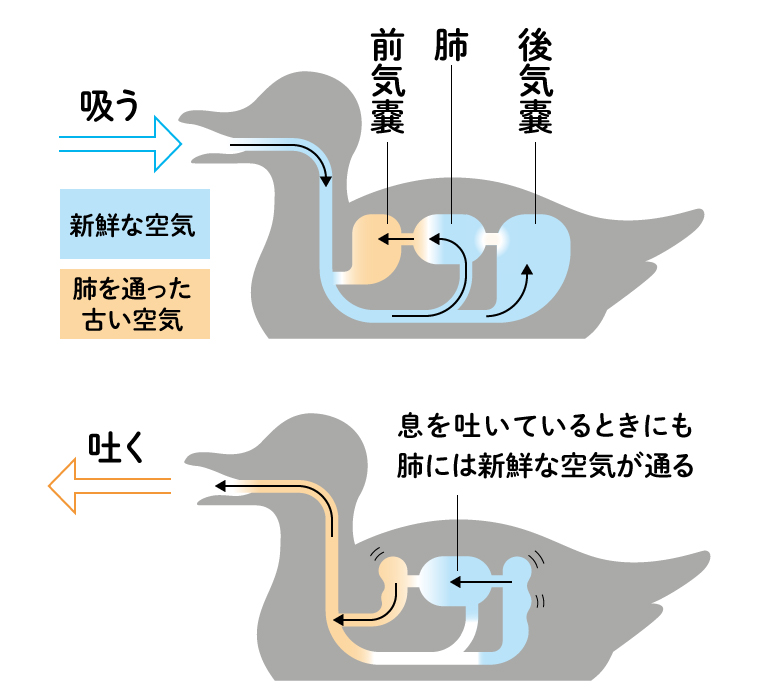
哺乳類の肺呼吸では、吸う息と吐く息が肺で混じりあってしまいますが、気嚢システムでは、肺にはいつも気嚢から新鮮な空気が送られます。
7.哺乳類のゲノム
昨今技術が進み、哺乳類の何十億塩基という長さのゲノムDNAの並び方も機械に任せて決めることができます。しかし、機械から出てくる配列は短く分かれていて、ゲノムの中によく似た配列が繰り返し見つかるので、つなげるのには苦労します。ジグソーパズルのよく似たピースを思い浮かべましょう。その似た配列は転移因子といって、ゲノムの中で増えたり、移動したりする邪魔ものですが、ヒトのゲノムでは半分近くが転移因子です。転移因子の中には、ウイルスとよく似た配列もあり、かつて感染したウイルスが入り込んだようです。ところが、この転移因子が、いつの間にか、大切な役割の遺伝子になっていることがわかりました。哺乳類の特徴である、お腹の中で子供を育てる胎盤は、ウイルス遺伝子を拝借してつくるようになりました。神経の伝達に関わるシナプスのタンパク質やお肌の角質化など、さまざまなところで転移因子が由来の遺伝子がはたらいていることがわかってきました。ウイルスのような邪魔者もうまくとりいれて、ゲノムはしなやかに、したたかに、進化してきたのです。カンブリアから続いてきたしくみを守り、組み合わせや使い方を変えながら工夫するのも、外からやってきたものも受け入れる柔軟さもどちらもゲノムの特徴です。
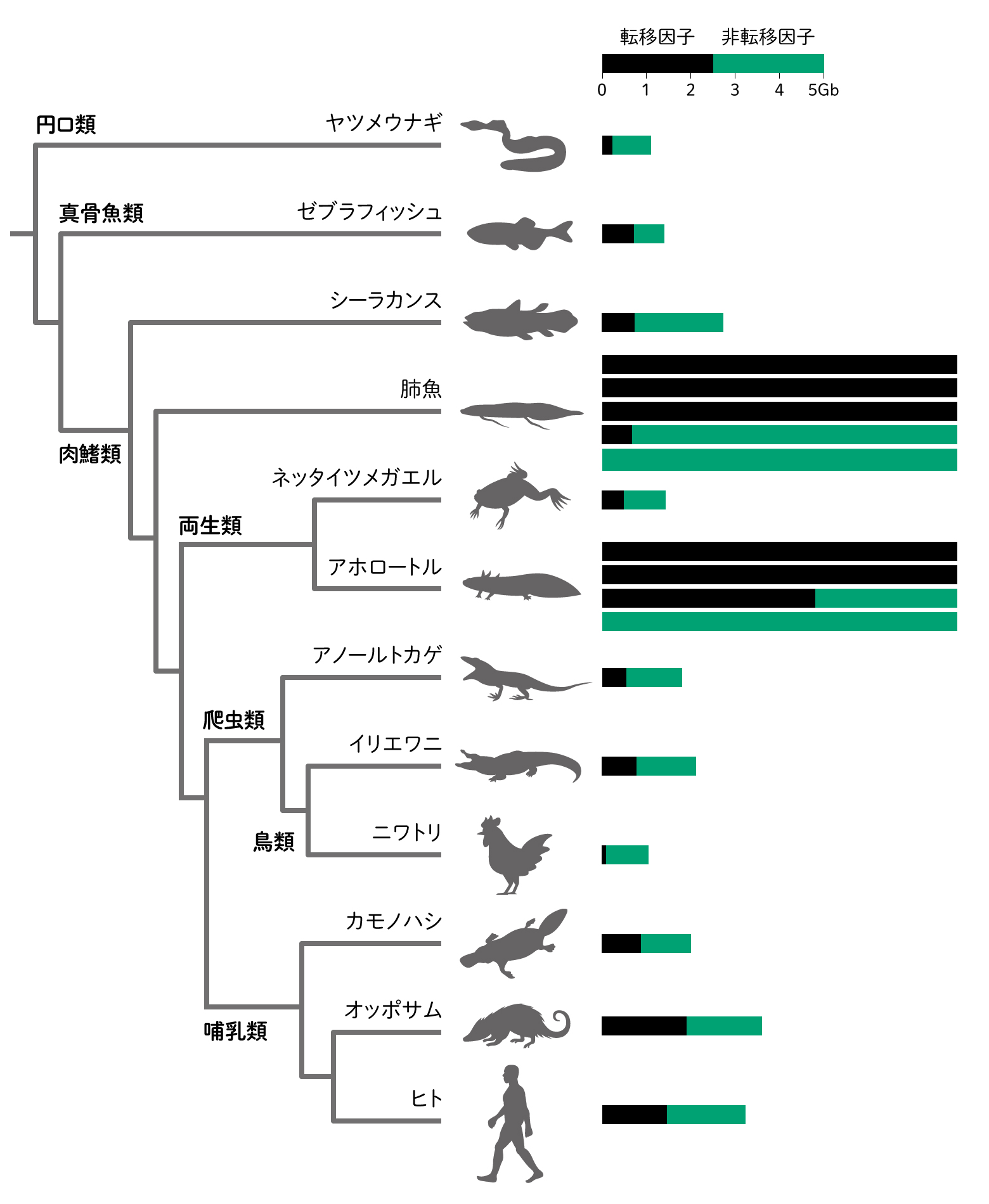
(図7) ゲノムサイズと転移因子
棒グラフはゲノムの大きさとその中で転移因子が占める割合を示します。さまざまな細胞で異なる遺伝子がはたらくことにも転移因子が関わっています。
文献[11]のFigure2より改変。
8.生命誌のゲノム
生きもののゲノムは、親から子へと伝わり続いてきましたが、時には、他の生きものからの配列を取り入れ、組み変わり、変化して、環境に適応したさまざまな生きものを生み出し、豊かな生態系に育まれてきました。時には厳しい地球の変化を乗り切って、生き残ってきたのが今のわたしたち生きものの世界です。ゲノムには、困難を克服したしかけや、ともにあった生きものとの関わりが刻まれているのです。ところが現在、人間の活動が第6の大量絶滅を引き起こしていると言われます。しかし、これまでの大絶滅で消えていったのは、その時代の生態系を象徴するものであったことを思い出してください。これからも生命誌の物語を語り継ぐために、人間は生きものであり、自然の一部であると、あらためて考える時です。

参考文献
[1] Colnaghi M et al. Repeat sequences limit the effectiveness of lateral gene transfer and favored the evolution of meiotic sex in early eukaryotes. Proc Natl Acad Sci U S A. 119(35):e2205041119 (2022)
[2] Ocaña-Pallarès E, M et al. Divergent genomic trajectories predate the origin of animals and fungi. Nature 609(7928):747-753 (2022)
[3] Yin Zet al. Developmental biology of Helicoforamina reveals holozoan affinity, cryptic diversity, and adaptation to heterogeneous environments in the early Ediacaran Weng'an biota (Doushantuo Formation, South China). Sci Adv. 6(24):eabb0083 (2020)
[4] Simakov O et al. Deeply conserved synteny and the evolution of metazoan chromosomes. Sci Adv. 8(5):eabi5884 (2022)
[5] Parker HJ, et al. An atlas of anterior hox gene expression in the embryonic sea lamprey head: Hox-code evolution in vertebrates. Dev Biol. 53(1):19-33 (2019)
[6] Bi X et al. Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes. Cell. 184(5):1377-1391.e14 (2021)
[7] Griffin DK et al. Dinosaurs: Comparative Cytogenomics of Their Reptile Cousins and Avian Descendants. Animals (Basel). 13(1):106 (2022)
[8] Uno Y et al. Inference of the protokaryotypes of amniotes and tetrapods and the evolutionary processes of microchromosomes from comparative gene mapping. PLoS One. 7(12):e53027 (2012)
[9] Rens W et al.The multiple sex chromosomes of platypus and echidna are not completely identical and several share homology with the avian Z. Genome Biol. 8(11):R243 (2007)
[10] Pereira, Núbia P., et al. "Karyotype characterization and nucleolar organizer regions of marsupial species (Didelphidae) from areas of Cerrado and Atlantic Forest in Brazil." Genetics and Molecular Biology 31: 887-892 (2008)
[11] Sotero-Caio CG et al.. Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol Evol. 9(1):161-177 (2017)
文責:平川美夏

JT生命誌研究館 表現を通して生きものを考えるセクター
研究成果やそこに至る考えをどのように表現するか、独創的な表現は新しい考えを生み出す源泉です。セクターの紹介はこちらへ。
.jpg)

.jpg)
.jpg)










