

Experiment
ミドリムシは動物?植物?
分子系統樹が藻類の世界に迫る
葉緑体をもちながら動物のように動くミドリムシは、はたして植物なのか、動物なのか。
高等植物は緑藻に属すシャジクモの仲間から進化してきた、と考えられているが、これは本当だろうか。
生命誌研究館が遺伝子を使って明らかにした藻類の進化の一断面--。
CHAPTER
食卓に並ぶコンブやアサクサノリは褐藻(かっそう)や紅藻(こうそう)と呼ばれ、身近な藻類の一例である。一方、かたい殻をもち鞭毛で泳ぎ回る渦鞭毛藻や、ケイ酸質の殻をもち池や海に浮遊するケイ藻は、単細胞の生き物であるが、同じく藻類の仲間に分類される。藻類といっても、細胞の体制や色、生活の仕方はさまざまであり、1つの仲間として把握することは難しい。一般に、原生生物のうち、水中で生活し光合成をする生物のことを藻類と呼ぶ。
どの藻類と、どの藻類が近縁なのか。これは、藻類全体の系統進化を考えるうえで重要な問いであるが、形態や生活史(どんな一生を送るか)などの比較だけでは十分な結論は得られない。私たちは、特定の遺伝子の変化に着目して分子系統樹を作成し、藻類の進化の歴史を明らかにすることを試みた。
解けたミドリムシの秘密
ミドリムシ(ユーグレナ)は、葉緑体をもって炭酸同化作用を行い、植物のようにでんぷん類を栄養として蓄えることができる。しかし、植物にある細胞壁がなく、先端についている鞭毛で泳ぎながら、体をくねくねと変形させる。また、光のない環境では、外部から食物を取り込んで生きていける動物的な要素を備えている。このため、植物学者はミドリムシ植物門のミドリムシ藻類に、一方動物学者は原生動物門のミドリムシ類に分類するというありさまで、いまだに動物か植物かの決着がついていない。
私たちがミトコンドリアの遺伝子(COX I)を使って、藻類以外の生き物(原生動物や菌類)のデータとも比較した結果では、ミドリムシは原生動物の仲間に近いと出ている。ほかの研究グループによる核の遺伝子を用いた解析でも、鞭毛虫(水や、動物の消化管などに棲む50ミクロンほどの微生物)に近い原生動物の仲間と出て、私たちの結果と同じである。しかし、葉緑体が独自にもっている遺伝子による解析では、緑藻の仲間と出ている。この一見不思議な結果は、細胞同士の「共生」で説明できる、というのが私たちの結論である。
共生による多重膜の形成の模式図(仮説)
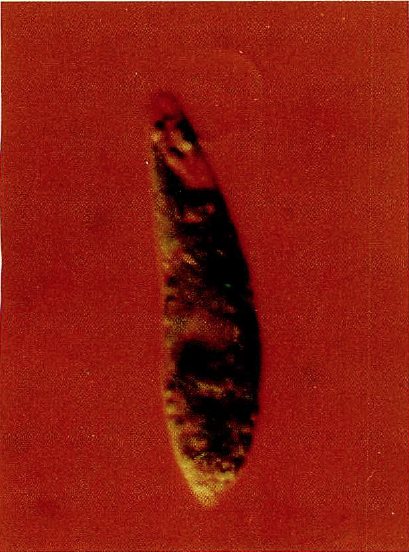
ミドリムシ。三重膜の葉緑体をもつ。
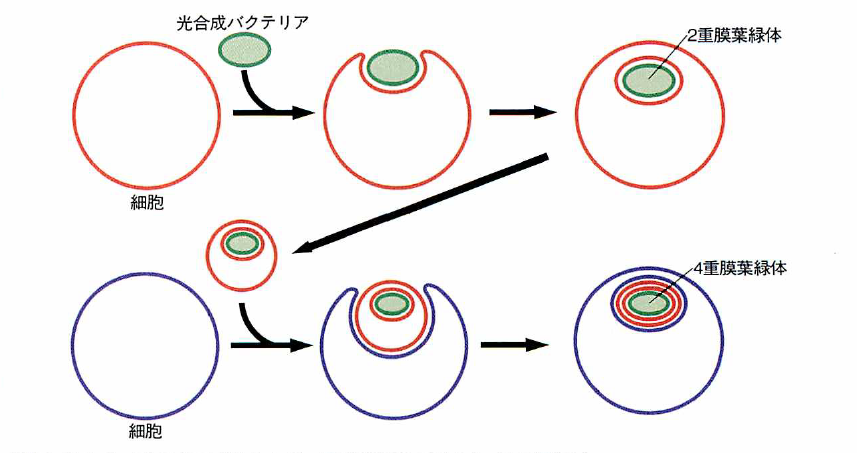
細胞に光合成バクテリアが共生して二重膜葉緑体ができる。この細胞がもう1度ほかの細胞に共生し、四重膜をもつ葉緑体が生じる。
緑藻の仲間や高等植物は、葉緑体が二重の膜で包まれている。これに対して、ミドリムシの葉緑体は三重膜である。いろいろ調べてみると、緑藻などの一部の仲間を除いた藻類には、三重や四重の膜をもったものがかなりある。しかも、四重膜をもつクリプト藻では、退化したと思われるもう1つの核(核様体=ヌクレオモルフ)が見つかっている。こうした事実は、二重膜の葉緑体をもつ単細胞生物がほかの単細胞生物と共生し、その後核の1つが消え、外来の葉緑体だけが四重膜となって残った、というできごとが進化の過程で起きたことを推測させる。ミドリムシの場合は、原生動物の細胞に藻類が入り込み、その後、藻類の核とミトコンドリアは退化して消失したが、葉緑体は残った、と私たちは考えている。葉緑体の膜は、一部で融合したために3枚になったのかもしれない。
この仮説なら、核とミトコンドリアの遺伝子が示す系統的位置づけと、葉緑体の遺伝子が示す系統的位置づけとが違っていることも、矛盾なく説明がつくのではないだろうか。おそらく、藻類が藻類に共生したり、藻類に原生動物が共生するなど、進化の過程では生物同士のダイナミックな相互乗り入れが頻繁に起きていたに違いない。そうしたことも、藻類の成り立ちを考えるうえで今後明らかにしていきたいと思っている。
進化は共生の歴史
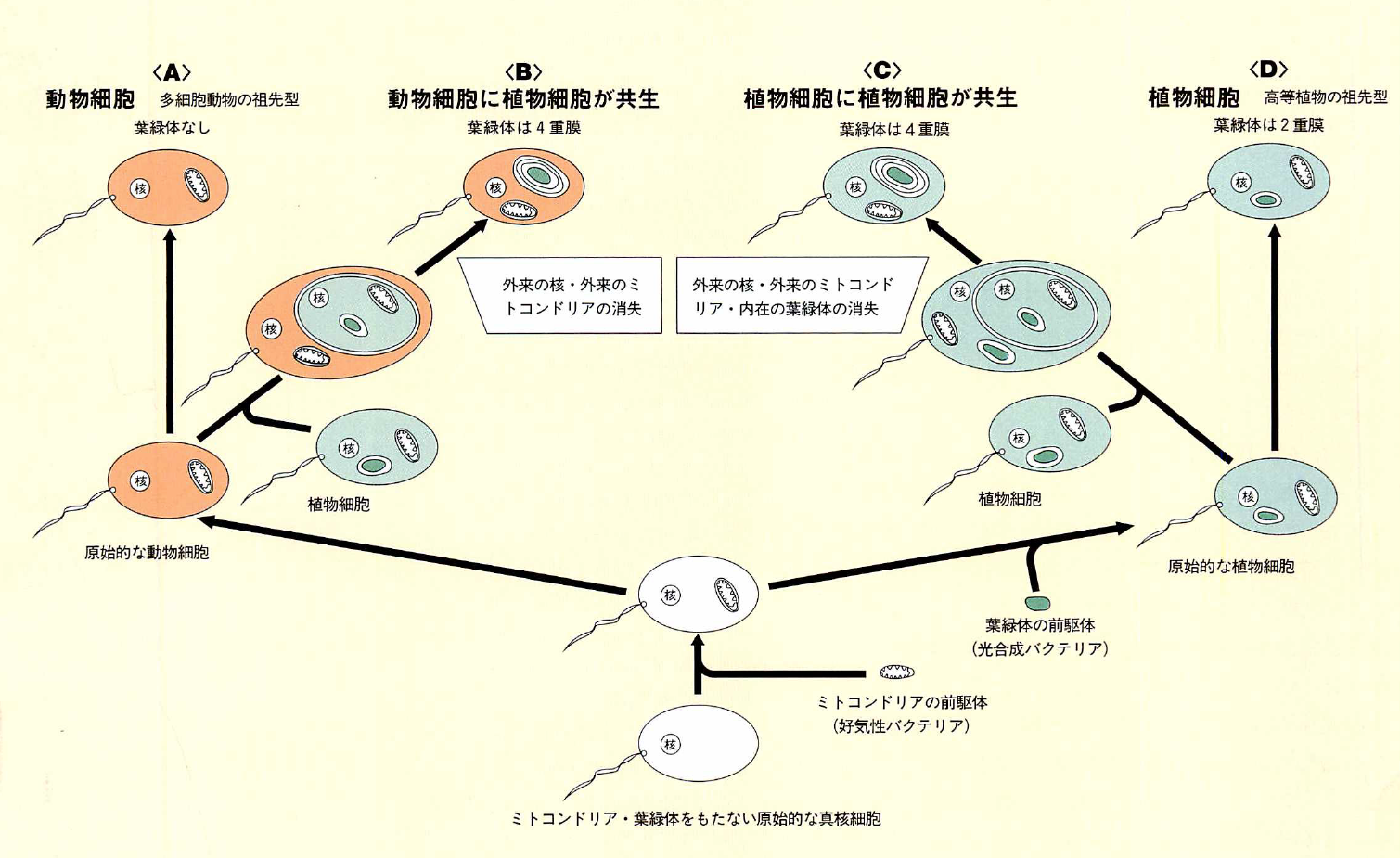
何回もの共生のステップをへて、多様な細胞形態が生まれた。ミドリムシは原始的な動物細胞に、さらに藻類(植物細胞)が共生したものだと考えられる。
細胞同士の共生体
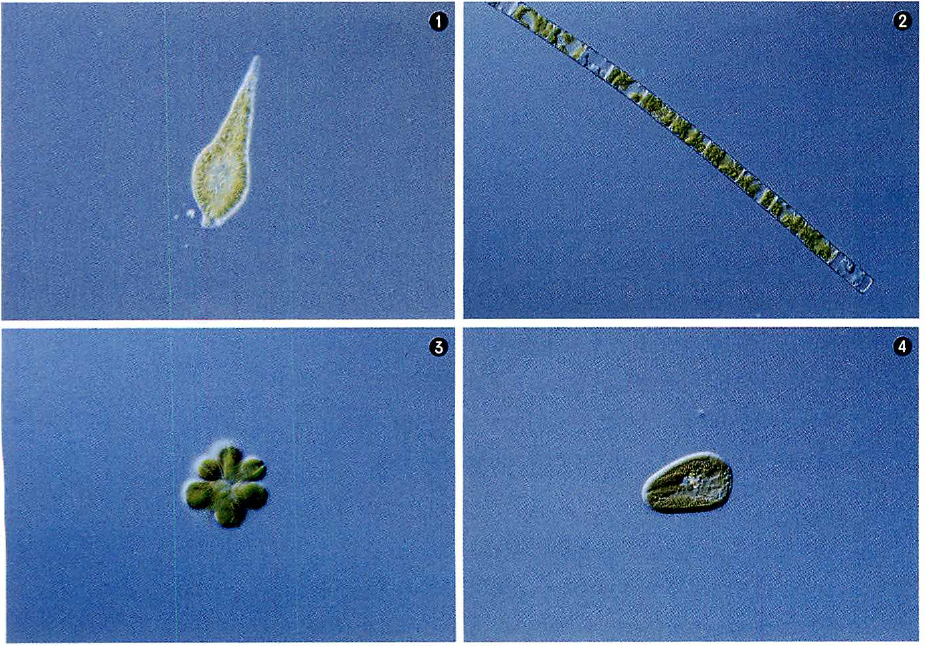
いずれも四重膜の葉緑体をもち、宿主となった細胞に植物細胞が共生して葉緑体をもち込んだことをうかがわせる。宿主が動物細胞(上図B)か植物細胞(同C)かは解析中。
①ラフィド藻 Chattonella antiqua
②ケイ藻 Melosira ambigua
③黄金色藻 Synura petersenii
④クリプト藻 Cryptomonas ovata
高等植物の起源
藻類は、鞭毛の構造や貯蔵物質の種類、光合成色素の種類などから14種類に分けられる。らん藻は光合成色素としてクロロフィルaしかもたない。陸上植物と同じクロロフィルa、bという2種類の光合成色素をもち、炭酸同化作用ででんぷんをつくる藻類の仲間は、私たちが日常よく見かけるサクラやバラといった高等植物の直接の祖先だと考えられている。
緑藻は、形も構造もじつにさまざまである。ミドリムシと同じく鞭毛をもち泳ぎ回るクラミドモナス、群体をつくることで知られるボルボックス(オオヒゲマワリ目)、鞭毛のない単細胞性のクロレラ、糸状に細胞が並ぶアオミドロ、5cmを超える巨大細胞のカサノリ、コケのように造卵器・造精器をつくる多細胞のシャジクモ(車軸藻)。
緑藻は、単純なものから複雑なものへと進化し、もっとも複雑な体制をもつシャジクモの仲間から高等陸上植物が分かれてきた、とするのが通説である。しかし、ミトコンドリアの遺伝子の遺伝子を使った私たちの系統解析では、シャジクモが高等植物の起源であるとの説を有力に裏づける結果は出なかった。シャジクモは高等植物よりも他の緑藻に近いと出ている。
今回の解析で、シャジクモはコレオケーテやツヅミモと同じグループに分けられた。このことは、鞭毛の微細構造や核分裂の様式などを比較した最近の研究報告と符合している。
緑藻が複雑な体制を進化させる以前の単細胞藻類だった時代に、陸上植物の祖先は緑藻からすでに分かれたのではないだろうか。その後独自に複雑な体制へと進化し、陸上生活に適応していった-そんなストーリーが考えられるのだが、どうだろうか。
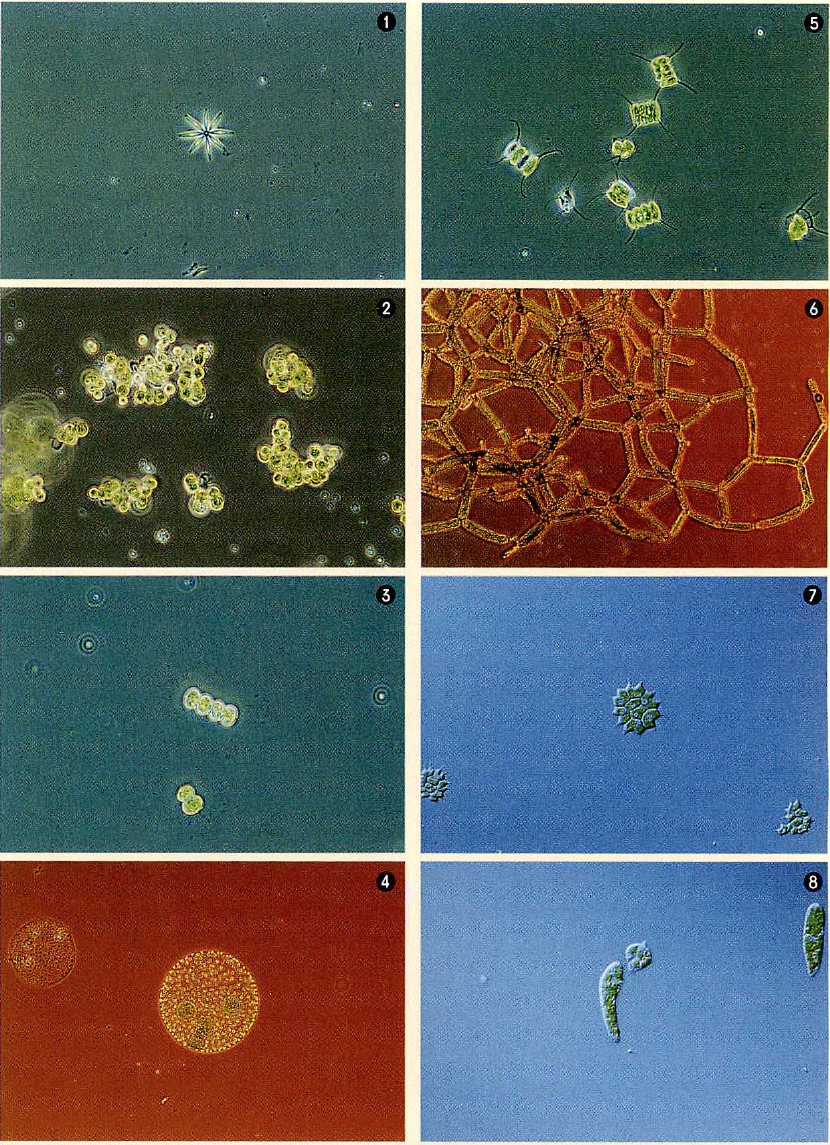
系統樹上のさまざまな藻類
①アクチナストルム。星のように美しい群体をつくる。
②クロレラ(Chlorella reisiglii)。球形の単細胞生物。
③ツヅミモ。ツヅミ状のものが1つの細胞。
④ボルボックス。群体の中に娘群体をつくる。
⑤イカダモ。普通4個の細胞でイカダ状の群体をつくる。
⑥アミミドロ。細長い細胞が網袋状の群体をつくる。
⑦クンショウモ。群体が勲章のように見える。
⑧ミドリムシ(Euglena gracilis)。丸いのも変形したミドリムシ。
多様化の歴史を探る
群体をつくる緑藻類の仲間が多細胞生物の起源ではないか、といわれたことがある。私たちの遺伝子解析によると、群体をつくるアミミドロの仲間は、ミトコンドリア遺伝子の遺伝暗号UAGが、アミノ酸のアラニンやロイシンに翻訳されるように使われている(下図「ミトコンドリア遺伝子の解析に基づく分子系統樹」※印)。しかし、高等植物では、この暗号は遺伝情報の読み取りストップを意味するもの(=終止コドン)として使われている。遺伝暗号の変化は、一度起こるとその子孫に受け継がれていく。つまり、アミミドロの仲間では遺伝暗号の変化が起きており、高等植物の進化の道筋からは離れたところにあることがわかる。
今回の解析で、遺伝暗号が変化しているものが緑藻の中の5つの種で見つかった。その多くが群体をつくることは興味深い。これらは細胞同士の結合がゆるいため糸状体にはなれず、進化の方向として、これ以上複雑になれなかったのだろう。このグループにのみ遺伝暗号の変化がみられることが、それを支持するのではないだろうか。遺伝情報の面から藻類の多様化の問題に迫った1つの例である。
これまで系統の位置づけがはっきりしなかった緑藻の仲間を、はっきりと位置づけることもできた。たとえば、アクチナストルムは群体性で、クロレラ目のなかでも同じく群体を形成するイカダモに近いと考えられていたが、イカダモよりも単細胞性のクロレラに近いといえる。また、ヒビミドロ目としてクロレラ目とは別に分類されていたヒビミドロ(Ulothrix zonata)が、思ったよりもクロレラ目と近縁であることがわかった。さらに、同じクロレラ属の2種(C.vulgarisとC.reisiglii)がそれほど近縁でないことや、単細胞性のテトラエドロンが群体性のアミミドロやクンショウモに近縁であることなども明らかになった。このように、分子系統学は、形態や構造などの特徴だけでは不明瞭だった藻類の分類法として有効であることが示されたのである。
ミトコンドリア遺伝子の解析に基づく分子系統樹
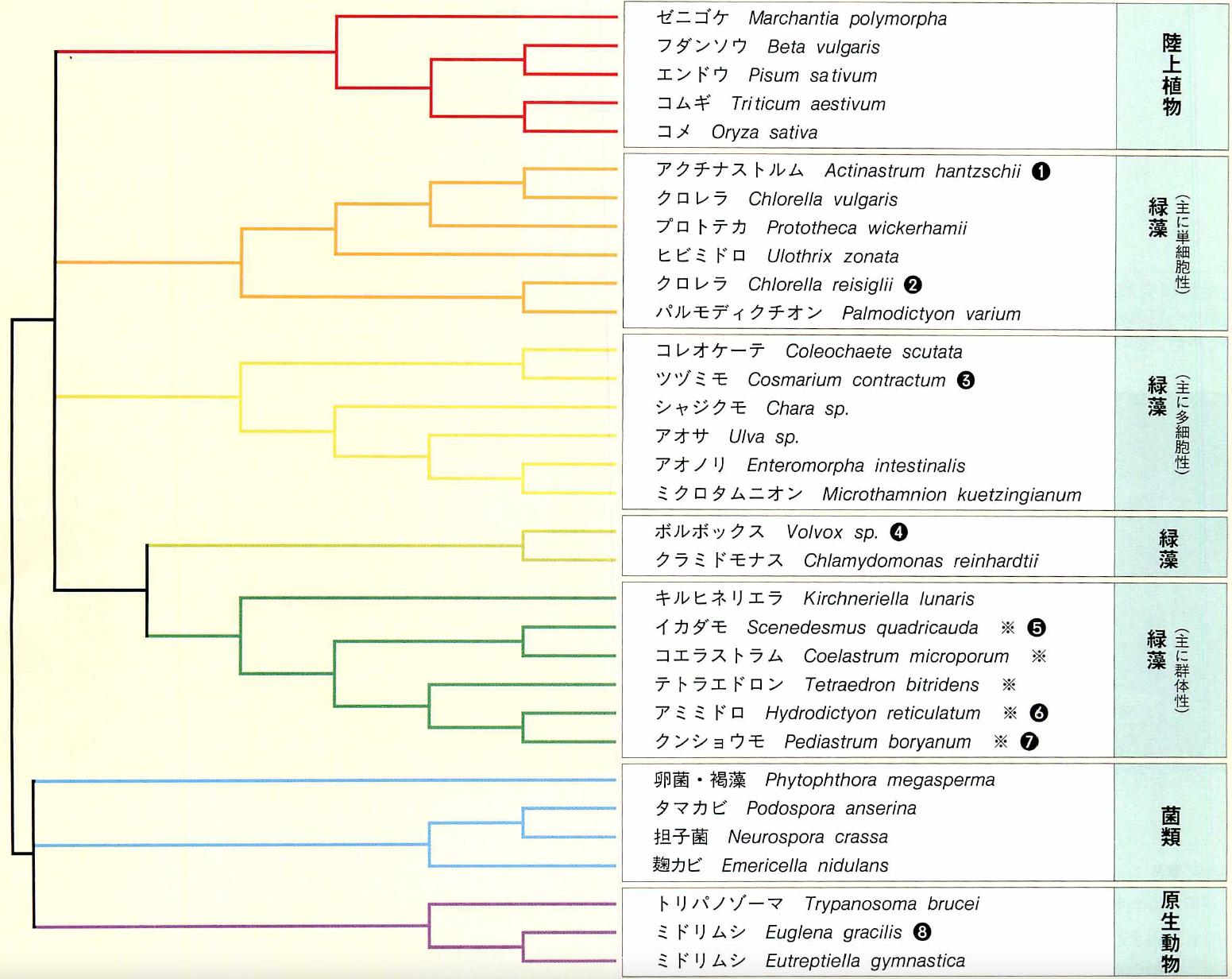
緑藻全体は単一の系統でなく、高等植物の起源もかなり古い可能性がある。シャジクモが高等植物に近いというデータは得られなかった。ミドリムシの仲間は他の原生動物や菌類と近縁で、緑藻の仲間との分岐は古くさかのぼることがわかる。(※印は遺伝暗号が変化した仲間)
(いしまる・やすこ/生命誌研究館奨励研究員:顕微鏡写真とも)
※所属などはすべて季刊「生命誌」掲載当時の情報です。.jpg)

.jpg)
.jpg)










