

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
システム化学
2016年1月15日
ここまでの話をもう一度ざっとまとめると、次のようになる。
熱水噴出孔ではエネルギーと炭酸ガス、水素、水など無機分子の持続的な流れが発生している。この流れはそのままだと、海水へと放出されるだけだが、噴出孔のマイクロチェンバーにより様々な形で外的に制約されると、有機分子の生成が始まる。最初の有機物はアセトンやメタンだが、マイクロセルの成分に含まれる様々な金属の触媒が作用すると、さらに複雑で多様な有機物が合成される。例えば以前紹介したように(第42回ゲノムの発生学II)、モンモリロン石が存在すればアミノ酸や核酸の重合が進む。脂肪酸が選択的に合成されるマイクロセルも存在するはずだ。たまたま存在した様々な外的制約により、分子やエネルギー分布の不均一化、構造化がおこる。
このようにマイクロセル内で進む有機分子の持続的な合成の流れを用いて、さらに複雑な有機高分子の合成が始まるが、ここまでのほぼ全ての過程はマイクロセルによる制約があれば進む。こうしてできた様々な分子(低分子から高分子まで)は独立したまま存在していることもあるが、高分子になるほど低いアフィニティーであっても他の分子と相互作用する確率が高まる。すなわち、合成されてきた分子自体が他の分子と反応する能力が新しい制約として働き出す。システムの中で生まれた新しい分子が、システムを新たに制約する「再帰的」制約の発生だ。
この「内的に発生する再帰的制約」について、これまでも具体的例を紹介してきた。例えば、現存するAutotroph細菌を見ると、アセトンを原料にするのは真正細菌のclostridia、メタンを原料にするのは古細菌と完全に分かれ、逆はない(第43回熱水噴出孔と生命誕生)。即ち、最初に合成された成分が制約要因としてLUCA発生から現在まで、持続的制約としてAutotrophで働き続けていることを意味する。他にも脂肪酸の中には核酸の重合や複製を触媒する能力がある分子が存在することを紹介したが(第42回ゲノムの発生学II)、これも内的に生まれた分子が、システムの新たな制約要因として働き、それ以前には存在しなかった新しい分子を作り出す分子進化の例といえるだろう。
この内的に生まれた制約による新しい状態の誕生は、制約要因が最初のシステムには全く存在しなかったという点で、反応する溶液同士を混ぜ合わせた系でおこる振動、ジャボチンスキー反応とは違う。システム内で発生した分子が制約となって、それまでとは異なる秩序が生まれるのが「内的に発生する再帰的制約」だ。
これは机上の空論だと思われるかもしれないが、応用有機化学分野のひとつ、コンビナトリアル化学では同じような課題が現実に研究されている。コンビナトリアル化学とは、例えば創薬のための候補化合物を、単純な分子から一挙に作ってしまおうという技術で、様々な化学工業で利用され始めている。従来多数の化合物を得るためには、自然に存在する生物を集め、そこから化合物ライブラリーを調整するのが普通だったが、コンビナトリアル合成のおかげで、生物に頼らず試験管の中だけで多様な化合物を合成することが可能になっている。
オランダのOttoらの総説(Li et al, J.American Chemical Society, 135:9222, 2013)は、この技術により単純な有機化合物からこれまで存在しなかった様々な新しい機能を持つ化合物が合成できることを教えてくれる(図1)。総説の内容を詳しく紹介するのは控えるが、単純な有機化合物がコンビナトリアル合成により、無限の可能性を拓くことがよくわかる。
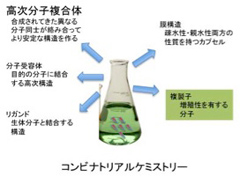
図1 コンビナトリアルケミストリーの可能性を述べたOttoらの総説の内容を図示した。有機分子が反応を続けることで様々な複雑な高分子を合成できることを示している。
中でも私の興味を引いたのが、コンビナトリアル化学から自己複製する分子構造(複製子)が生成される可能性だ。この複製子について、この総説の著者であるオランダのSijbren Ottoのグループが昨年Nature Communicationsに発表した論文(Colomb-Delsuc M et al, 6: 7427, 2015 Creative Commons)に基づいて紹介しよう。
まずこの実験では、グリシン・ロイシン・リジン・フェニルアラニン・リジンの疎水性と親水性のアミノ酸が互い違いに繰り返すペプチドが、互いに可逆的共有結合を形成できる(SS結合)SH基を2つ持つベンゼン環に結合したオリゴペプチドが使われた(図2)
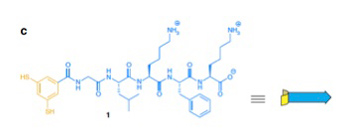
図2 Ottoらの実験では、図に示す構造を持つオリゴペプチド溶液を攪拌するだけの実験が行われた。
このオリゴペプチドはベンゼン環部分に2つあるSH基を介して他の分子と反応し、図3に示すような1merから7merの重合体を形成し、最終的な平衡状態に達すると予想される(図3)。
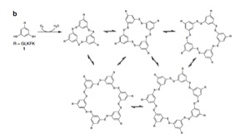
図3:図2に示したオリゴペプチド同士の反応から予想される分子同士の結合反応。相互に反応するベンゼン核のみを示している。
実際には7merは合成されなかったようだが、攪拌を続けると、4日目ぐらいまでに1mer,2mer,3mer,4mer,そして6merが混ざった平衡状態が生まれる(図4)。ところが、1週間を過ぎた頃から急速に他の重合体は失われ、6merが全体の90%を占めるようになる。
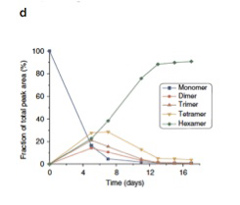
図4:16日間攪拌を続けた間に溶液中に存在する各重合体の比の変化。
5日目に予想された平衡状態に達するが、この平衡は1週間目に破れ、6merが急速に優位になることがわかる。
なぜ最初の平衡が破れたのかを調べると、6merの大きさの重合体だけが、今度は縦に重なって、さらに大きな重合体を形成していることが明らかになった(図5)。即ち、6merだけが縦に重なって成長する能力をたまたま持っていたため、一旦この6merが発生すると、それが核となって、6merが重合した高次構造がうまれる。この高次構造は成長に伴い、溶液中から6merは隔離するため、平衡状態が壊れる。その結果、溶液中には6mer由来の巨大分子だけが残される。
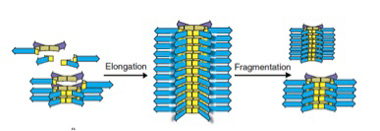
図5:6merは重なって縦に伸びる能力を持つ。この結果、溶液は6merによって占められるようになる。この重層構造は一定の大きさに達すると物理的力で壊れるが、6merの供給が続く限り、別れた部分から元の大きさの構造が形成される。すなわち、複製が起こる。
この一連の反応は、6merという構造単位の発生が新たな制約になり、これまでとは全く異なる高次構造を形成することが現実に起こりうることを示している。面白いことに、6merが縦に重なった重合体は、一定の大きさに達すると、攪拌のストレスにより、自然にちぎれる。もし6merが持続的に供給されれば、ちぎれた部分がもう一度核になって、同じ大きさを持った重合物が再形成される。この意味で、この分子構造は複製能力を持っていると言える。
このような重合による高分子形成は、ウイルス被膜や、細胞外マトリックス形成など、生命過程では普通に見られる自己組織化反応だが、全く人工的な反応として取り出してみると、生命と共通の幾つかの性質が生まれていることに気づくことができる。
- 1) 核酸情報は持たないが、特定の構造の複製が可能なこと。
- 2) 複製のためには、外界から部分が常に供給される必要があること(同化)。
- 3) 部分を構成する分子組成は全く同じでも、重合することで外界とは隔離された系(すなわち自己)が形成されていること。
- 4) 同じものを作るためには情報が必要なこと。ただこの場合の複製に必要な情報は、外界のストレスと、構成分子の性質(この場合は6merしか受け入れない科学的性質と、割れるまでの構造の強度)として提示されている。
これを流れの中の渦に例えると、力と水分子の定常的な供給が続く、流れの中に同じ大きさと形をした渦が増え続けるのと同じことだ。
先週Ottoたちはこの構造にグリシン・ロイシン・リジン・セリン・リジンを持つオリゴペプチドも同化できることを発見し、両方のオリゴペプチドが存在する溶液では多様な構造体が生まれることを示した(Sadownik et al, Nature Chemistryオンライン版)。この結果は、この分子構造が上の4つの性質に加えて、進化するという5番目の生命と共通の性質を持ちうることを示している。もちろん多様化だけでは進化は起こらず、特定の形質が選択されることが必要だが、このモデル系は、自然選択による進化とは何かのヒント教えてくれそうだ。
コンビナトリアル化学では最初の反応条件は人間が決める。熱水噴出孔で初期条件を決めているのは偶然だが、様々なオリゴペプチドが形成されることは既に確認されており、Ottoたちが観察したのと同じことが起こって何の不思議はない。次に、Ottoたちの研究から生命と共通するどんな性質が生まれてくるのか楽しみだ。
Otto等の研究を詳しく紹介してすこし長くなったので、今回はここまでで終わる。次回はDeaconの説を紹介しながら、生命に共通の性質の発生のためのルールについてさらに考えてみたいと思っている。