

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
情報の進化II クロマチン構造を媒体とした情報
2016年9月1日
フェロモンの次に議論する情報は「クロマチン構造を媒体とした情報」だ。この情報は一般的にエピジェネティックスと呼ばれることが多い。ただ、エピジェネティックスという言葉の指す範囲は曖昧で広い。このため、ここではエピジェネティックスを「クロマチン構造を媒体とした情報」に限定する。
まず簡単に「クロマチン構造を媒体とした情報」、エピジェネティックスについて簡単に解説する。
私たち人間は40兆個近くの細胞を持っているが、全て一個の受精卵が発生過程で、発生プログラムに従って様々な細胞に分化した結果だ。リンパ球のような一部の例外を除くと、何百、何千種類もの異なる細胞ができるためにゲノムの変化は全く必要ない。細胞の形は異なり、作られる分子は全く違っていても、私たちの体の中の細胞は全て同じゲノムを持っている。このゲノムの変化なしに異なる性質の細胞が発生し維持されるメカニズムがエピジェネティックスだ(図1)。
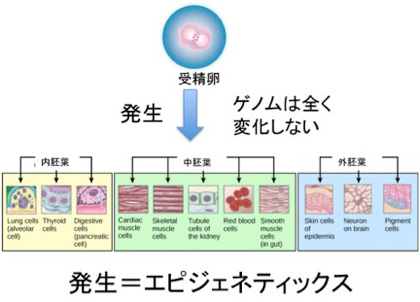
図1 発生ではゲノムは変化せず、クロマチン構造の変化だけが起こる。従って、ほとんどの発生過程は、エピジェネティックな過程と見ることができる。(図はWikiを拝借)
次にこの発生の結果何が可能になったか考えてみよう。何百種類の細胞がただ発生したというわけではない。例えば皮膚の細胞は皮膚細胞を作り続け、血液細胞は血液を安定に作り続けることが可能になっている。同じゲノムを持っている細胞同士なのに、決して皮膚の細胞が血液に変わることはない。もしそんなことが一定の確率で起こったら私たちの体の統合性はたちどころに破綻する。
これを支えるメカニズムがクロマチン構造を媒体とした情報=エピジェネティックスで、原則として真核生物で初めて見ることができる。このゲノムとヒストンと呼ばれるタンパク質が互いに協調することで形成されるのが、クロマチン構造で、それぞれの細胞種でパターンが異なる。また、細胞分裂でヒストンが外れても、もう一度同じように再構成できることから、クロマチン構造を繰り返し形成し直すための一種の鋳型ができていると考えると判りやすい(図2)。
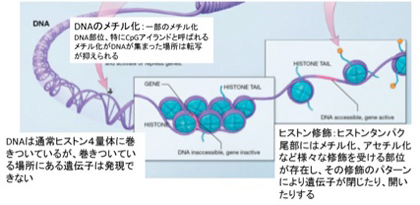
図2:クロマチン構造と修飾:ヒストンの少ないところはクロマチンが開いていると表現し、一般的にそこにある遺伝子は転写因子と相互作用する。一方、多くのヒストンが集まった部位はクロマチンが閉じていると表現し、ここにある遺伝子は転写因子から隔離されており、発現できない。(図はWiki を拝借)
このクロマチン鋳型は、全ゲノムにわたって活性化部位と、不活性化部位を区別する印をつけることにより形成される。これらの印は、ノートの開きたい場所と、閉じておきたい場所につける付箋のようなものと考えるといい。
体の全ての細胞には、それが属する種類、すなわち皮膚細胞、血液細胞といった種類に対応する異なる鋳型が存在している。そして何よりも重要なのは、細胞分裂でこの鋳型も複製され、二つの娘細胞に伝わる。このような性質から考えれば、エピジェネティックスをクロマチン構造を媒体とした情報と呼ぶことは何の問題もない。また、この鋳型があるおかげで皮膚細胞は間違って血液細胞や脳細胞になる心配なく、皮膚細胞を作り続けることができる。実際、この鋳型を維持するメカニズムが狂うと、皮膚細胞が思いもかけない細胞へ分化するといった乱れが生ずる。ガンで見られる異型細胞の出現はこの例だ。
ではこのクロマチン鋳型とは何で、情報としてどのような役割を担っているのか見てみよう。すでに図2に図示したように、ゲノムは細胞の種類に応じてヒストンが詰まった閉じた状態と、結合しているヒストンが少ない開いた状態にわかれている。
この開いた状態、閉じた状態を決めているのが、染色体と結合しているヒストンのN末端にある領域に存在する数種類のリジン残基のメチル化とアセチル化、そしてDNA自体のメチル化だ。このゲノムの場所特異的なヒストン修飾パターンやメチル化DNAパターンが鋳型として子孫に伝わり、繰り返し働く。これを可能にするのが、発生過程で細胞分化に合わせて鋳型を書き込むメカニズムと、細胞分化のあと完成した鋳型を維持するメカニズムだ。
このメカニズムについての理解は最近急速に深まってきたとはいえ、今も研究途上の分野なので、ここでは解説は省く。ただ、多能性幹細胞を始め、様々な培養細胞のクロマチン鋳型を精密に調べる研究から、
- 1)細胞が分裂を繰り返しても同じ鋳型が子孫細胞に受け継がれること、
- 2)多能性幹細胞から様々な細胞系列が分化する過程で、各細胞に特異的なクロマチン鋳型が形成されること、
- 3)分化細胞をそのまま培養しても普通はクロマチン鋳型が変化することはないが、iPS誘導のように特殊な条件下ではリプログラムが可能で、この時実際に変化するのはこのクロマチン鋳型であること。
などが明らかになった。
この結果は、発生や成長で細胞分化のシグナルに応じてクロマチン鋳型を書き換えるメカニズムと、書き換えたクロマチン鋳型を安定に維持するメカニズムが間違いなく存在することを示している。すなわち、塩基配列のようなコード性は存在しないが、クロマチンのパターンを解釈して複製できることから、エピジェネティックスをフェロモンの次に誕生した情報と呼ぶことができる。
最後にクロマチン構造を媒体とした情報の特徴を、ゲノム、フェロモンと比較しながら見てみよう。
前回見たように、フェロモンはリアルタイムの個体間のコミュニケーションを可能にした全く新しい情報で、ゲノムだけがどれほど複雑化してもなしえない可能性を拓いた。前回このフェロモンの役割について、単位あたりの個体数を感知して一定の反応を起こすquorum sensingを例に説明した。ただフェロモンが活躍するのはquorum sensingだけではない。研究が進むもう一つの領域が、バクテリア同士がプラスミドを受け渡すときに働くフェロモンだ。これによって薬剤耐性が多くのバクテリアに伝搬するため、医学にとっては重要な分野になっている。
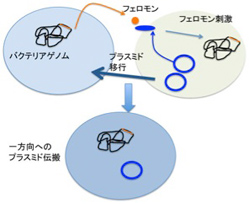
図3:フェロモン刺激によりプラスミドを持つバクテリアから持たないバクテリアへのプラスミドの伝搬が起こる。このメカニズムで薬剤耐性遺伝子が伝播される例が多く知られており、医学でも問題になっている。
図3に示す腸球菌の系では、フェロモンはバクテリアのゲノムにコードされているが、その受容体は薬剤耐性を持つプラスミドにコードされている。この結果、フェロモンに反応するプラスミド(+)バクテリアと、フェロモンを分泌するプラスミド(ー)バクテリア同士が特異的な接合を起こし、プラスミド陰性のバクテリアにプラスミドを渡すことが可能になっている。実際には、プラスミド(+)バクテリアが自分のフェロモンに反応しないよう、うまく抑制システムができているが、詳細は省く。
この現象からわかるのは、フェロモンが引き金となって、DNAを媒体とする情報のやりとりが行われ、この結果、効果が一過性のフェロモンのシグナルが、遺伝子変化として記録されることだ。すなわち、一過性のフェロモンの効果を長期間保持するためには、バクテリアでは遺伝子の変化として記録する必要がある。しかし、プラスミドのような限られた情報の伝搬は簡単でも、ゲノムを書き換えたりすることは難しく、出芽酵母のような一部の例外を除いて、同じ書き換えを繰り返すことは難しい。
一方真核生物への進化により、一過性に誘導される遺伝子発現状態をもとに、クロマチン鋳型を形成する能力、すなわちエピジェネティック過程が誕生すると、遺伝子の書き換えなしに、外界からのシグナルを特定のクロマチン構造として書き残すことが可能になる。
これをバクテリアがゲノムに記録する場合と比べると、外来の情報を持続的に記録するという点では同じだが、
- 1)記録が書き換え可能
- 2)多くのシグナルに対応して記録が可能。
- 3)同時に多くの個体がシグナルを共有し、それを同じ記録として維持することが可能。
などの、ゲノムでは不可能とは言えなくとも、簡単ではない記録が可能になっている。
これがクロマチン構造を媒体とする情報の誕生により生まれた可能性で、この獲得には20億年、生物の進化全過程の半分以上の時間が費やされている。次回は、この新しい情報の進化と、この情報がもたらしたインパクトについて考える。