

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
外界の情報を記憶するためのエピジェネティックス
2016年9月27日
前回「クロマチン構造変化を媒介とする情報(<エピジェネティックス)」が、
1)情報の書き換えができる、2)様々な外界のシグナルの影響を比較的迅速に記憶できる、3)異なる個体間で同じ情報を共有して同じ反応を可能にする、の3点で、ゲノムを媒体とする情報と異なることについて述べた。今回はこの違いについてもうすこし具体的に掘り下げる。
これを読んでいる読者のほとんどは、ルイセンコの名前を聞いても誰かわからないだろう。ルイセンコは、メンデル遺伝理論だけでは説明がつかなかった植物の現象を捉えて、獲得形質は遺伝することを主張したソビエトの農学者だ。彼を政治的に支持したスターリン政府の後ろ盾を得て、論敵を逮捕・追放したことで、科学的議論に政治を介入させたとして科学史に大きな汚点を残した人物だ。彼の理論は我が国にも様々な影響を残したようだが、これは私が大学に入学するより前の話だ。幸い、ルイセンコと彼の理論をめぐる我が国での議論については本館名誉顧問の岡田節人先生が生命誌ジャーナルに紹介されているのでぜひ読んでほしい。
さてルイセンコがメンデル遺伝学の反証の根拠にしたのが「春化(春処理)」と言われる現象で、植物を低い温度に晒すことで開花時期が調節できるという現象だ。低温という環境で獲得した形質が、世代を超えて伝わる場合もあることから、獲得形質が遺伝すると考えたようだ。結局エピジェネティックス(遺伝した後)の問題で、ゲノム自体の変異を基盤とする遺伝学の問題でないことが明らかになる。このように、春化メカニズムは、エピジェネティックス、すなわちクロマチンの構造を介した情報の問題として現在解明が進んでいる。外界からのシグナルがクロマチン構造を媒体とする情報として書き換えられるメカニズムを理解するのにうってつけの例なので、まずこの春化とそのメカニズムについて見てみよう。
各々の植物は発生過程で開花を調節する機構を確立する。これには温度や日照時間を感知しながら様々な遺伝子の発現を調節する複雑なメカニズムが関与する。話をわかりやすくするために、実験のためのモデル植物として研究されているシロイヌナズナの開花を抑制するマスター遺伝子FLCに絞って春化の現象を解説する。
まず越冬一年草型シロイヌナズナのライフサイクルを見てみよう。秋に発芽、ロゼット型葉を形成してゆっくり成長しながら越冬する。その後、茎が伸びて開花、そして夏になると種ができる。この間の開花抑制遺伝子FLCの発現を見てみると、発芽時の秋には最も高いレベルにある。しかし、ロゼットが形成される冬には急速にそのレベルが低下し、春の開花期には0になる。開花が終わると夏に向けて急速に発現レベルが回復する。
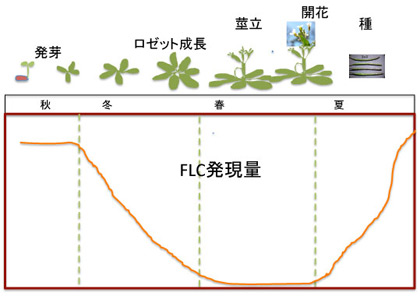
図1 シロイヌナズナの開花と、開花抑制遺伝子FLCの発現量。説明は本文参照。花と種の写真はWikipediaより。
FLCが開花抑制に関わることを頭において発現の季節パターンを見ると、開花の起こる春以外は開花抑制遺伝子であるFLCを発現させ、間違った時期に開花が進まないようできているのがわかる。すなわち、秋から冬にかけて気温が下がり始まるのを合図に、FLCの発現を安定に抑制するための準備を始め、この状態で急速に温度が上がると、開花を優先した植物の発生が進み、開花する。その後、種ができ、発芽するまでにFLCの発現を戻して開花を抑え、植物はエネルギーを成長に使うことが可能になっている。
次に、このパターンの背景にあるメカニズムを細かくみていこう。この季節に合わせた発現の調節にはFLC遺伝子領域とそこに結合しているヒストンのメチル化パターンが大きな役割を演じている。すなわちFLCの発現を抑えるときはこの領域に巻きついているヒストン3のN末27番目のリジンがメチル化されており(H3K27me3)、一方発現が高いときは4番目のリジンがメチル化されている(H3K4me3)(図2)。すなわち、クロマチンが閉じて転写が抑制されるか、あるいはクロマチンが開いて転写が行えるかは、ヒストンの標識の違いによって決まる。
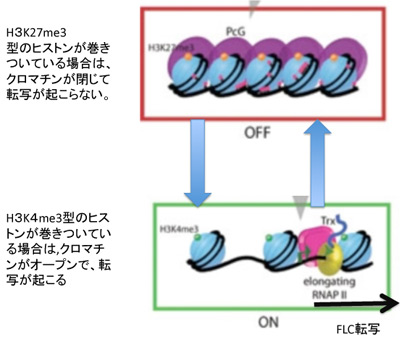
図2 ヒストンのメチル化によるクロマチンの構造のスウィッチ。図はES細胞分化時に起こるクロマチン構造変化についての説明をStem Bookサイトより転載。(http://www.stembook.org/node/585.html)
つまり、FLCに巻きついているヒストンのメチル化の異なる2つの状態を温度スウィッチでシフトできるようになっている。このようにクロマチン構造を媒体とする情報の誕生は、外界の変化を細胞自体の記憶に変えることを可能にした。
このヒストン標識によるクロマチン構造のスウィッチのメカニズムについてはずいぶんわかってきたが、複雑でその全像について解説するには本1冊必要になる。とりあえず、図にあるように、クロマチンの構造を維持する鍵になる分子が、ポリコム複合体とTrithorax複合体であることを理解しておけば十分だろう。
H3K4のメチル化と、H3K27のメチル化パターンを転換させクロマチン構造を変えることで大きな領域の遺伝子発現を調節するメカニズムは、実は酵母から人間まで保存されているメカニズムだ。事実、ポリコム複合体やTrithorax複合体という名前はショウジョウバエ突然変異の研究が由来となっている。例えば、ショウジョウバエから哺乳動物まで、体の前後軸に合わせて体節の特異性を決めているHox遺伝子が集まる大きなゲノム領域のどこまでを活性化し、どこまでを不活化するかは、まさにこのクロマチン構造変化により調節維持されている。
図3は哺乳類の発生でのHox遺伝子領域のヒストン標識パターンを図示しているが、Hox遺伝子の発現はクロマチン構造の変化で調節され、植物とほぼ同じメカニズムが使われていることがわかる(図3)
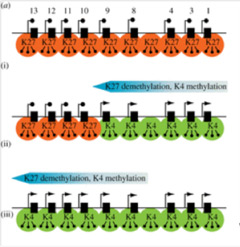
図3 クロマチン構造変化によるマウスHoxa1-13遺伝子の発現調節。内部細胞塊のような多能性幹細胞ではHox領域は完全にH3K27me3ヒストンで標識され、発現が抑えられている。前から後ろの体節に移行するのに合わせ(i)(ⅱ)(ⅲ)、Hoxa1からH3K4me3ヒストンは拡大していく。
出典(http://rstb.royalsocietypublishing.org/content/368/1620/20120367)
最後に、春化時のクロマチン構造の変化の際、他生物で見られない特徴について見ていこう。実は、FLC遺伝子発現を抑制するH3K27me3標識の範囲の量を調節できるようにできている。すなわち寒い時期が長いと、H3K27me3が結合するFLC遺伝子領域が拡大し、暖かくなってもこの抑制標識を外しにくくなる。これは、低温に長くさらされた植物の場合、開花が早くなり、開花期間は長くなり、さらに開花量も上昇するという量的な変化が、FLC遺伝子に結合するH3K27me3の量的な変化と相関していることを意味している。このように、外界からの情報を細胞内に量的な記憶として残すことができるのも、クロマチン構造を媒体とする情報の特徴だ。これをゲノム自体の調節機構で実現することはほぼ不可能だ。
春化を例に、クロマチン構造を媒体とする情報が、外界の情報を、細胞自体の情報に書き換えて記憶することを可能にしていることを示した。重要なことは、この記憶が書き換え可能で、量的にも変化させることが可能なことだ。進化が環境の自己への取り込みであるという観点から見ると、クロマチン構造を媒体とする情報は、ゲノムよりはるかに迅速に環境の自己への取り込みを可能にした新たな情報記憶の登場といえる。
今回例として使った春化では、環境の温度がクロマチン構造の情報として書き換えられる。一方、クロマチン構造の書き換えはフェロモンによっても誘導することができ、生命進化で誕生した新しい2種類の情報がここで出会う。すでに述べたように、フェロモンは個体間、細胞間のコミュニケーションに関わる情報と言える。一方、今回見てきたようにクロマチン構造を媒体とする情報は記憶媒体と言える。このように、コミュニケーションと記憶が揃った時、生命進化が大きく進む。こうして可能になったのが多細胞動物だ。次回はこの進化過程について考えてみる。