

RESEARCH
形づくりを支える
柔軟な遺伝子発現
多細胞生物の発生では、一つの受精卵が分裂を繰り返し、分化を経て多様な細胞が生み出される。それにより細胞はそれぞれ固有の機能を発揮するようになり、集団として組織・器官を構築し、それぞれ特有の形を持つようになる。胚発生の過程で、細胞が固有の機能を獲得することと、組織・器官として特有の形に変形することは同時に進行するが、両者はどのように足並みをそろえているのだろう。
1.機能と形の調和からなる組織
膨大な数の遺伝子と細胞が織りなす複雑な「発生」という現象により、どの生きものもそれぞれの形をつくり出す(動画1)。このような複雑な現象が正確に進行することは驚くべきことだが、実際に生きものはそれを成し遂げており、その背後には何らかのしくみがあるはずだ。個体発生は卵子と精子の受精にはじまり、分裂によって細胞の数が増えていき、各細胞は周囲から送られるシグナルや環境の変化によって特定の遺伝子発現が起こった後、それぞれ異なる種類の細胞へと分化する。一方、発生過程では細胞は遺伝子発現が起こると同時に、各細胞の振る舞いが協調することで自己組織化的にそれぞれ異なる形を持つ器官を構築していく(形態形成)。各細胞が遺伝子発現によって獲得する「機能」と、全体としての器官の「形」の両方がうまく同調することによって、器官は適切な生理機能を発揮できる。では、発生過程において同時進行する「遺伝子発現」と「形態形成」という二つの現象は、どのように調和するのだろか?発生過程では「遺伝子-細胞-組織」といった別々の階層をつなぎ、全体としての正確な発生の進行を保障する原理があると考えられるが、その詳細はほとんどわかっていない。
提供:吉氷康矢、近藤寿人
提供:京都大学 近藤武史
提供:JT生命誌研究館 小田広樹
提供:JT生命誌研究館 橋本主税
(動画1)さまざまな生きものの胚発生
2.ショウジョウバエの気管(trachea)形成
従来、発生過程では、まず細胞が周辺の分泌因子の濃度などに反応し、遺伝子発現により運命を与えられた細胞集団が、その遺伝子に従って特定の形をつくりあげていく、という考えが一般的であった(図1)。しかし、この一方向性の制御だけで本当に膨大な数の遺伝子と細胞が関わる発生・器官形成が正確に進行するのだろうか?
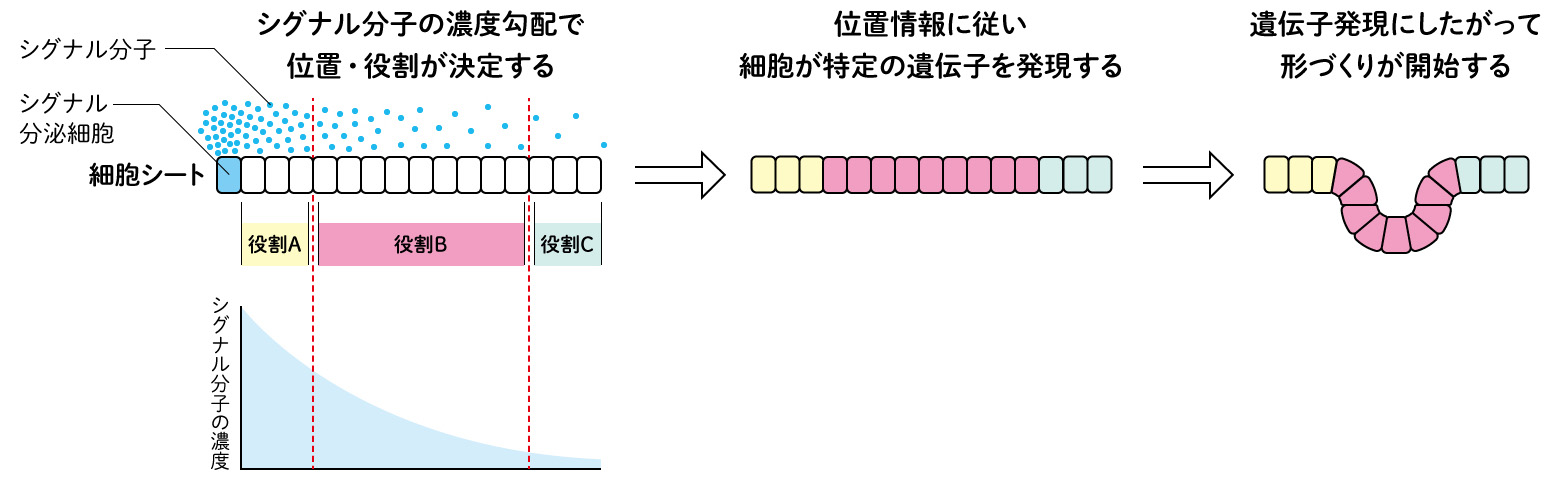
(図1) これまでの発生の考え方
形態形成の1例として、ショウジョウバエの胚発生で見られる空気を取り込む器官である「気管」の形成をモデルに研究を進めた。ここでは、シートに並んだ細胞が、体内に管状に陥入する過程に注目する。ショウジョウバエは、胚の発生中期に胚の外表面に一対ずつ、計10対の気管原基領域が誘導される(図2)。各気管原基はおよそ60個の細胞で構成されており、それぞれが陥入運動により二次元的なシート構造から三次元的な管状の構造へと変形する。その後、管状の構造が、分岐と融合を経て管ネットワークとなり体中に酸素を運ぶ。脊椎動物での血管のような器官である。
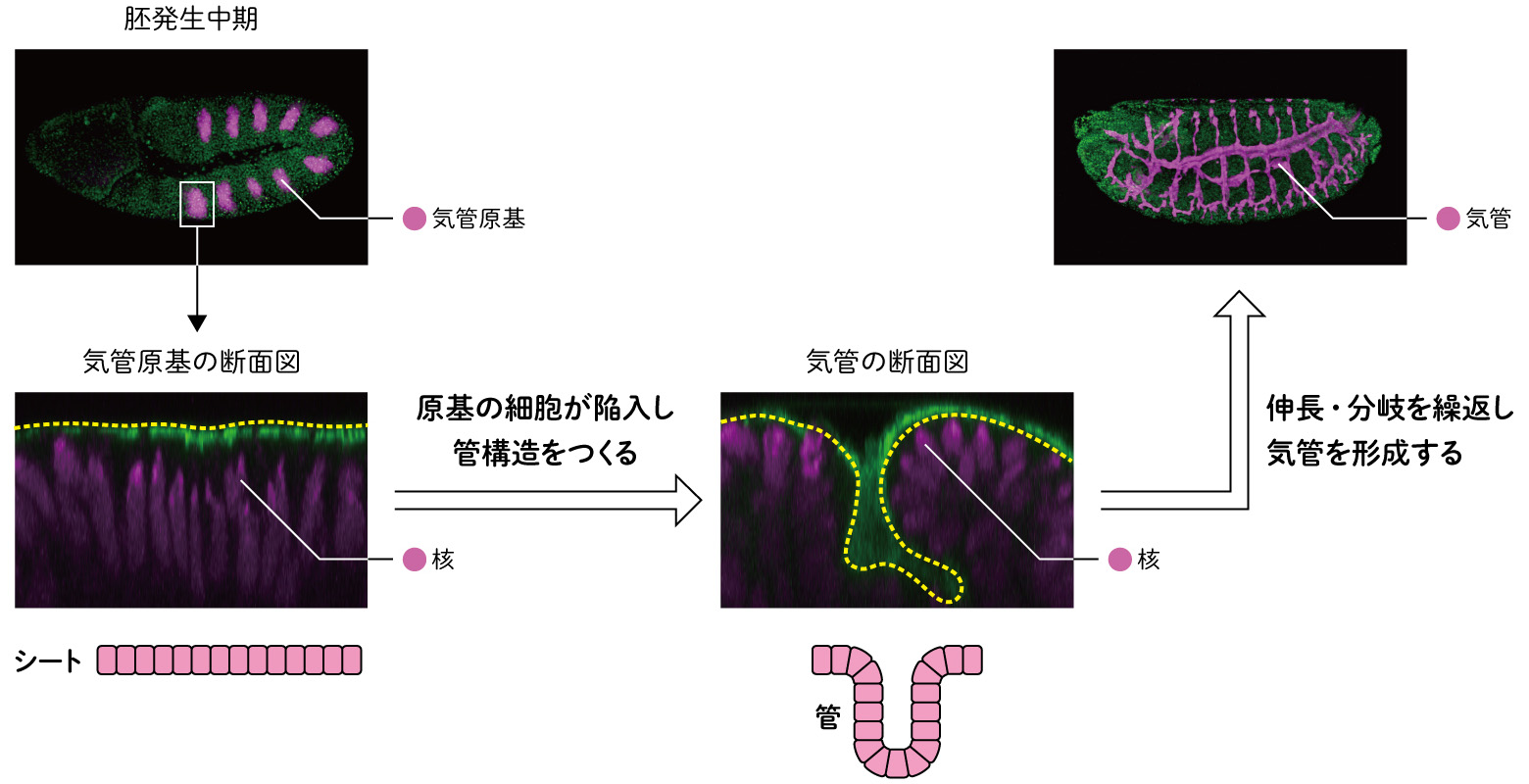
(図2) ショウジョウバエの気管形成
ショウジョウバエ気管細胞への分化を制御する遺伝子としてtrachealess(trh)という遺伝子が知られている。trhは転写を制御するタンパク質をつくり、その下流で様々な遺伝子の発現を制御することで気管細胞への分化を司っていると考えられている。私たちはこのtrhの発現を指標として、気管組織の発生過程を解析した。
3.trhを発現する細胞の運命
我々はこれまでの研究の中で、気管原基が誘導されtrhの発現は見られるが、その後の陥入がうまく進まない異常胚をいくつか同定していた。ここで同定した陥入が不完全な異常胚と正常胚を比較したところ、陥入前の気管原基においてtrhを発現する細胞(Trh+細胞)の数には大きな違いはなかった。一方、陥入後の細胞を調べたところ、trhは管構造を形成した細胞でのみ発現が観察され、陥入異常胚は正常胚に比べて陥入したTrh+細胞の数が減少していることが分かった(図3)。
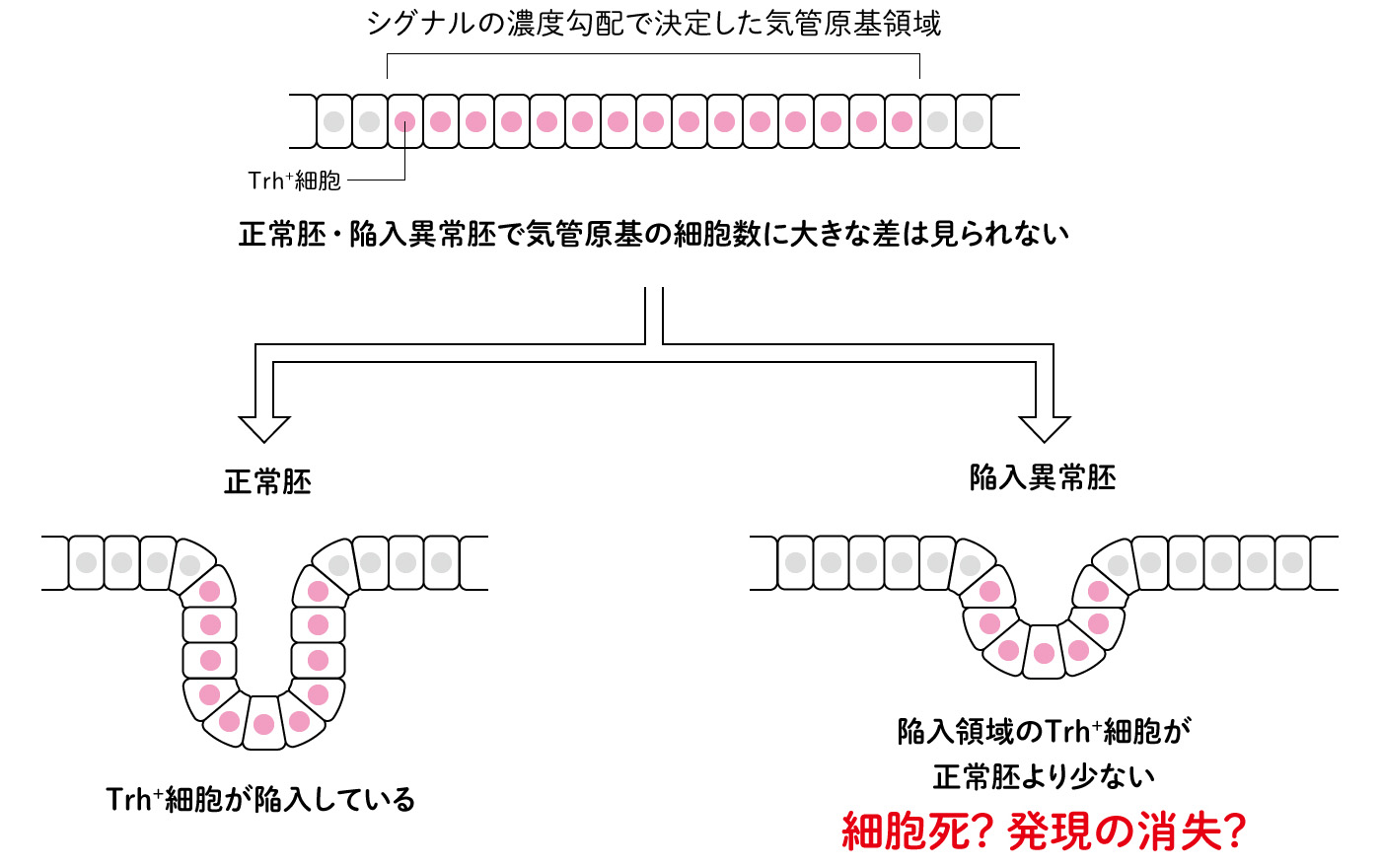
(図3) 陥入異常胚では陥入後Trh+細胞の数が減少していた
Kondo T, Hayashi S. Two-step regulation of trachealess ensures tight coupling of cell fate with morphogenesis in the Drosophila trachea. eLife. 2019 8:e45145. Fig. 2より改変
>論文サイト
遺伝子の変異体では、Trh+細胞はどのように失われたのだろうか。そこで、陥入前のTrh+細胞を標識し、その行く末を追跡した。まず正常胚を見たところ、驚くことに、Trh+細胞の一部が表皮に留まっており、すべてが陥入に参加していなかった。さらに、管の形成に参加しない細胞はTrh発現を消失した(図4)。一方、陥入異常胚では、正常胚よりも多くの細胞が表皮に留まり、それらの細胞もやはり後にTrh発現を消失していた。つまりTrh+細胞は、遺伝子の発現によって定められた運命に従って気管を形成するわけではないことがわかった。そして細胞は自身が陥入したかを認識することで気管細胞か表皮細胞がどちらの方向に進むのかを決定している、つまり遺伝子の発現には「形ができていること」が大切だと考えられた。
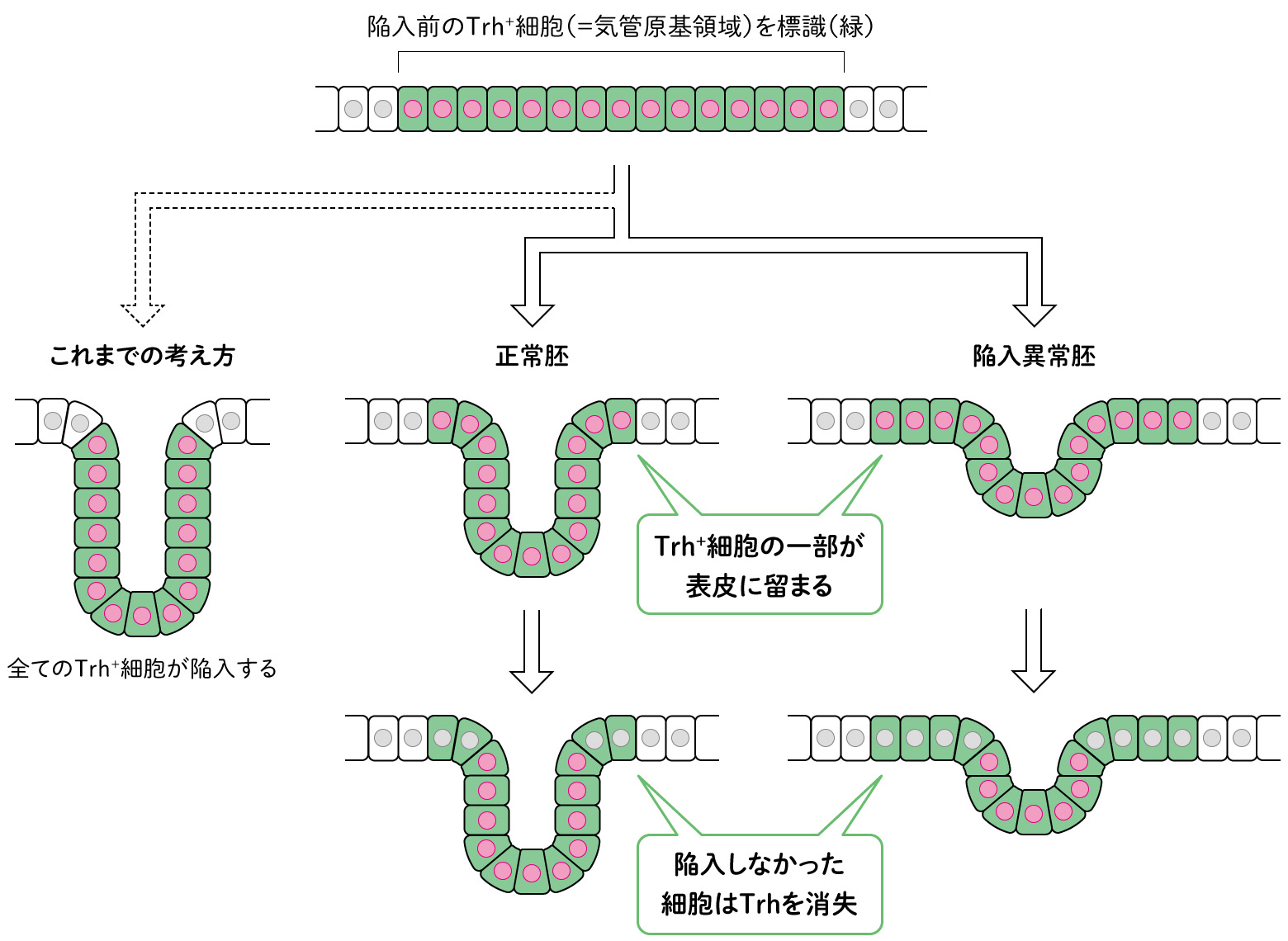
(図4)組織の形状に依存したtrhの発現調節
Kondo and Hayashi, eLife, (2019)Fig. 2より改変
4.組織の変形と維持
trh遺伝子の変異体では、trachealess(気管がない)という名前からも想像できるように(ショウジョウバエではその表現型から遺伝子の名前が決められることが多い)、気管が形成されない。そのためtrh遺伝子は気管原基の陥入に必須な遺伝子であると考えられていた。しかし、実際に確認されてたわけではなかったので、ライブイメージング技術により、trh変異体の気管原基の細胞の動きをリアルタイムで観察した。すると、trh変異体でも気管原基領域の細胞が内側へと陥入する様子が見られた(図5)。では、なぜtrh変異体では気管が形成されないのだろうか。さらに観察を続けると、細胞は一度陥入を始めるが、管構造を維持することができず、シート状の構造へと戻っていたのだ。つまり、trhがはたらかなくても、陥入運動を開始することはできるが、陥入後に管構造を維持するためには、trhが必要だということになる。形態形成において、「組織の形態形成を進める」ことと、「変形後の形を維持する」ことは異なるしくみがはたらいているようだ。
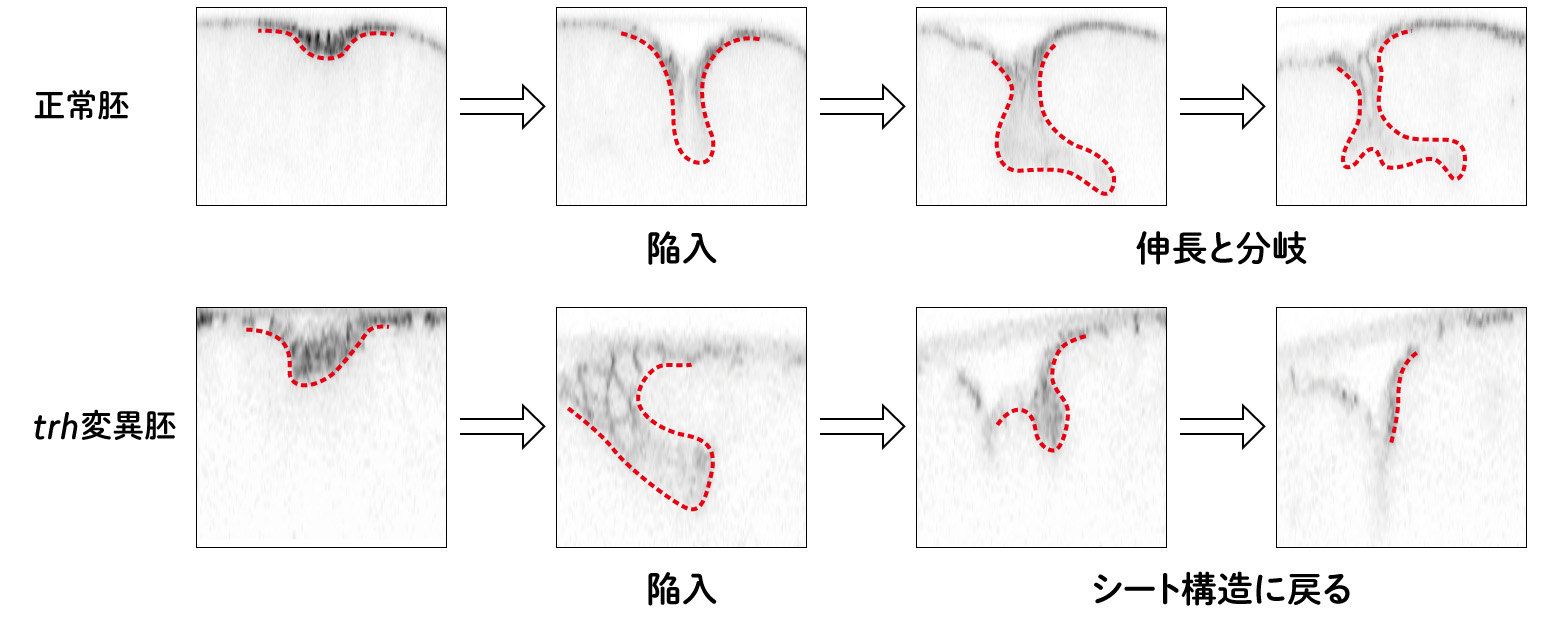
(図5) trhは、陥入後に構造を維持するために必要
Kondo and Hayashi, eLife, (2019)Fig. 1より改変
5.形態形成フィードバック
ショウジョウバエの気管形成の観察を通して、器官の形と機能の調和を保ち、正確な発生を進めるしくみが見えてきた。従来は「遺伝子発現」により細胞の運命が決まり、「形態形成」へ向かう一方向の制御と考えられてきたが、「形態形成」から「遺伝子発現」の逆方向の制御も存在し、その両方が必要だということである。ショウジョウバエの気管形成では、「陥入運動に伴う変形によりtrhの発現が安定し」、「trh遺伝子のはたらきにより構造が維持される」という、互いに調整を行うことによって、形と機能が調和した器官ができあがるのではないかと考えられる。
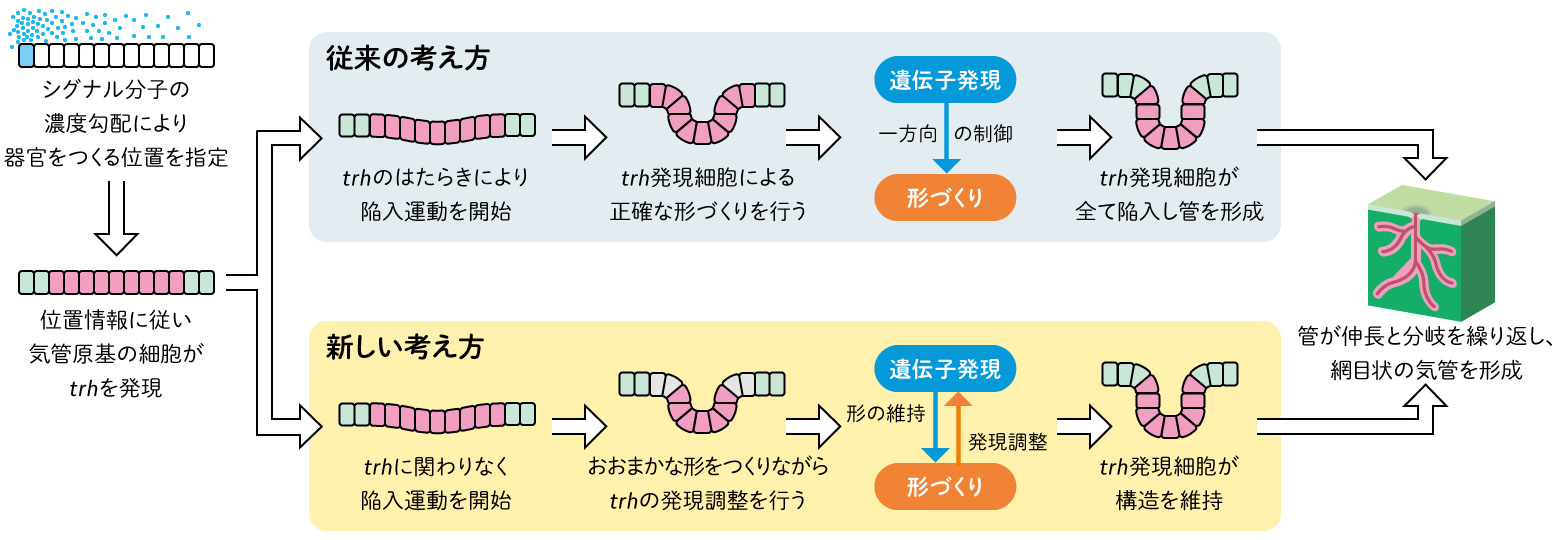
(図6) ショウジョウバエ気管形成における遺伝子発現と形態形成の調和のモデル
6.ばらつきのある集団から生まれる秩序
細胞の分化は遺伝子発現によって決まるが、その基盤は遺伝子の転写という生化学反応であり、オンとオフを繰り返す不連続なバースト状の反応ダイナミクスを示す(参考1)。また、組織・器官の形態形成を進める各細胞の振る舞いも詳細に観察すると、ゆらぎをともないながら変形している。つまり、多細胞器官を構成する要素(遺伝子・細胞)にはある程度のばらつきが存在する。にもかかわらず、全体レベル(器官・個体)では正確に発生が進行する(動画2)。ばらつきのある要素の集合であるにも関わらず全体としてはそのばらつきが積み重なって大きくなるわけではなく、むしろ秩序が生まれている。ここで我々が示した双方向の制御は、正確な発生を達成するための、基本的なしくみである可能性が十分にある。
提供:京都大学 近藤武史
提供:京都大学 近藤武史
(動画2)ばらつきのある細胞集団から生まれる秩序
7.これから
この課題に取り組むためには、イメージングによる観察に加えて、細胞分化の理解も深め、この独立した機構が同時に進行する関係を明らかにしていく必要がある。これまで細胞分化の解析では特定の遺伝子の発現を指標とすることが多く、分化が進行している最中の動態を捉えることが困難であった。しかし近年の大きな技術進歩、1細胞ごとの全遺伝子の発現を解析する1細胞RNA-seqとデータ解析環境が整ったことにより、この壁が取り除かれつつある。実際に私たちはショウジョウバエ原腸期胚の詳細な1細胞遺伝子発現アトラスの構築に成功しており(参考2)、今後、この技術を基盤にして細胞分化の動態を詳細に描写し、胚発生の時間の進み方を遺伝子、細胞、組織など、さまざまな階層で詳細に「観察」することによって、発生が正確に進行するしくみを理解していきたいと考えている。
(参考2) Okochi et al., Nature Comm. 2021; Sakaguchi et al., bioRxiv 2021

近藤武史(こんどうたけふみ)
2008年奈良先端科学技術大学院大学 博士後期課程終了。博士(バイオサイエンス)。学術振興会特別研究員(DC2、PD)、理化学研究所・発生再生科学総合研究センター・研究員を経て、現在は京都大学大学院生命科学研究科・特定講師。
.jpg)

.jpg)
.jpg)










