

VOL.1 実験発生学とオーガナイザー
近藤寿人(JT生命誌研究館 顧問・表現ディレクター)
今年、JT生命誌研究館の表現セクターの活動テーマとして取り上げる「生きものの時間」を語る上で、私たち自身――動物の発生を理解することは欠かせません。その学術分野である発生生物学は、15年ほど前から静かな革命を進めています。第一に、下村脩博士が発見されたオワンクラゲの緑色蛍光タンパク質などを用いて、生きたままの胚の中で特定の細胞がどのように変化していくのかを時間を追って調べることができるようになったこと。第二に、色々な生きもののゲノム情報(したがってどのような遺伝子が揃っているのか)が明らかになって、発生の一つ一つの段階を、遺伝子の働きの効果として理解できるようになったこと。これらが静かな革命を支えています。
では、どのような革命がおきているのか?それをお話しするために、今回は、これまでの発生生物学がどのようにしてできたのかを見てみましょう。次回からは、発生のプロセスで実際に起きていることと、これまでの発生生物学で「常識」のように語られていたことの間の大きな食い違いについて、異なった視点からお話しすることにします。
実験発生学の始まり
現在の発生生物学は20世紀の初めが出発点です。双眼の実体顕微鏡が発明されて、動物胚に対して、一部の組織を除いたり、ほかの胚の別の場所に移植したりすることが可能になりました。好んで用いられたのは両生類――特にイモリの卵から発生する胚です。イモリの卵は直径が2mm程と大きく、それから発生する大きめの胚は手術がしやすかったのです。このような手術を胚に施して、1日から数日経った時に、手術された胚がどのように発生したのかを調べて、その間に起きたことを推測し、それをもとにして、正常の胚発生を進める機構をさらに推測するーーという「実験発生学;Experimental embryology」と呼ばれる方法が定着しました。当時としては最先端の科学だったのですが、その方法の問題点と誤謬を引き起こす危険性について、有名な「オーガナイザー」を例としてお話ししましょう。
オーガナイザーの不確実さ
実験発生学の旗手の一人であったHans SpemannのもとでHilde Mangoldが原腸陥入を始めているイモリ胚の原口背唇部を取り出し、別の胚の側面に移植したところ、その移植の場所に、体幹部から尾に至るまでの、第二の胚というべき一続きの組織がつくられました(Spemann & Mangold, 1924)。のちに移植方法や、原口背唇部の範囲や、発生ステージを変えたりして、第二の頭部を作ることもできました(この辺りのことに関する、Johannes Holtfreterが活躍した1930年代の研究は、様々な重要な問題提起を含んでいたのですが、それについては機会を改めます)。この現象から、Spemannは、原口背唇部をオーガナイザー(発生過程全体を組織化する、発生の主催者といった意味合いがある)と呼んだのですが、原口背唇部をオーガナイザーと呼ぶことには、当時から強い異論がありました。Spemann自身1938年の著作で「私は現象の目覚ましさを表現しただけで、オーガナイザー説を唱えているわけではない」[1]と述べ、Spemann門下の2巨匠であるJohannes HoltfreterとVictor Hamburgerが1955年までの両生類の胚操作の研究を網羅的にまとめた総説[2]では、彼らは「仮にオーガナイザーと呼ぶことにする」と注意深く記載しており、Holtfreterは「オーガナイザーは名前のつけ損ない(misnomer)」[3]と言い切り、Hamburgerも1988年の名著「実験発生学が遺したもの:ハンス・シュペーマンとオーガナイザー」[4] の中で、オーガナイザーという考え方は再検討すべきだということを慎重な表現の中で幾度も述べています。それほど「オーガナイザー」と(仮に)呼ばれた胚の小領域が実際に何をしているのかは不確かなものだったのです。
オーガナイザーと両生類胚の特殊性
それにもかかわらず、オーガナイザーの名前と仮想の機能は神格化され、どの教科書にも事実として記載され、世界中に広まりました。Spemannが「オーガナイザーの発見」によって、1935年にノーベル賞を受賞したことの副作用もあったのかもしれません。とはいえ、後で述べるように、オーガナイザーと呼びたくなるような性質を持った、ある大きさを持った領域を両生類胚から切り出すことはできるので、両生類胚での問題は小さいのです。多くの脊椎動物の胚は、卵が胚に成長する栄養源となる卵黄を胚の外の袋の中に持っていて、胚はその卵黄から栄養を吸収しつつ発生するのですが、代表的な両生類の胚は、それぞれの細胞自身の中に卵黄を持つ(特に植物極側と言われる部分で豊富)という特殊な構成を持っています。その結果、一般的な教科書で模範例のように描かれている両生類胚の原腸陥入は、脊椎動物の中では例外的な細胞群の配置のもとで進行しているのです。(レクチャー記録映像「いつもとは違う視点から、生きものとしてのヒトを眺めてみよう」をご覧ください)
オーガナイザーに期待された働き
ここでオーガナイザーに期待された働きをまとめておきましょう。
ー 胚細胞の大掛かりな移動(原腸陥入)が起きている領域があるとともに、別の胚(宿主胚)の他の場所に移植すると(例え不完全であったとしても)主に宿主胚の細胞から構成される2つ目の胚構造を生み出すという働きをするもの。
ー 2つ目の胚構造を構成する胚の部分としては、(1)頭部、(2)骨格や筋肉そのほかのもととしての中胚葉組織、(3)胴体の中心線に沿って伸びる、もう一つの中胚葉組織である脊索、(4)消化管のもとになる内胚葉組織などが作られなければなりません。
他の脊椎動物の場合
「オーガナイザー」が神格化された結果、『他の脊椎動物の胚の発生も「オーガナイザー」という中核的な組織の働きに依存して発生するはずである』と考えられ、そして、哺乳類と鳥類の胚ではノード(ヘンゼン結節とも呼ばれる)といわれる小領域、魚類胚では胚盾と呼ばれる不均一な組織集団が、実験に基づく根拠があやふやな状況で「オーガナイザー」であると決め付けられてしまったのです。そしてさらに、鳥類の場合として、次のような図がまことしやかに教科書に描かれるようにさえなりました。このような鳥類胚を用いたノード移植実験は、私たちが実行するまで行われたことはなかったし(実際には、胚の培養技術を改善して初めて可能になった難しい実験)、この図のような結果は決して得られることはないのです。科学からの逸脱と言わざるを得ません。
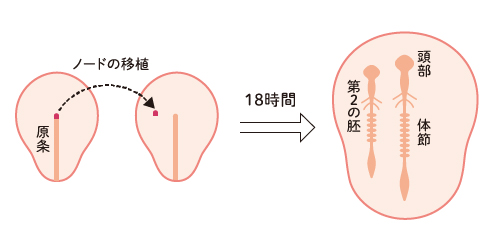
(図1)国際的なレベルの教科書に、まことしやかに書かれている想像 ※これは決して起きない
ノードは “オーガナイザー” なので、別の胚(宿主胚)のノード以外の場所に移植すると、宿主胚由来の組織からなる、頭部から胴部までを持つ二次胚を生み出す。
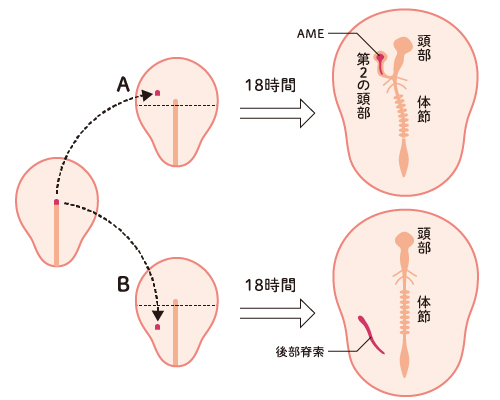
(図2)実際に起きること
A: ノードを胚の前側に移植すると、ノードから前側に伸び出した AME(赤)の周囲に、第二の頭部ができる。
AMEの移植でも同じことが起きる。ノードの直接の効果ではない。
B: ノードを胚の後側に移植すると、ノードから後部脊索が伸び出すのみで(赤)、第二の体幹部が作られることはない。
実験発生学のどこが問題だったか
先にお話ししたように、「実験発生学」は、「手術を胚に施して、1日から数日経った時に、手術された胚がどのように発生したのかを調べて、その間に起きたことを推測し、それをもとにして、正常の胚発生を進める機構をさらに推測する」ものでした。オーガナイザーの場合も、移植された原口背唇部はどのような細胞の組み合わせでできているのか?そのどの部分の細胞がどのように変化したり移動したりしてゆくのか?移植細胞に接する宿主胚の細胞がどのように変化するのか?といった情報が一切ない状態で、推測を積み重ねた、いわば砂上の楼閣のモデルだったのです。最初に述べたように、今では、生きたままの胚の中で、蛍光タンパク質で標識された個別の細胞がどのように変化していくのかを時間を追って調べることができますから、古典的な実験発生学に立脚したモデルは、もう一度ゼロから調べ直す必要があるのです。
私たちの実験 [5]
私たちは、脳を中心とした胚の頭部がどのようにしてできるかを調べるために、ニワトリ胚のエピブラスト(体の組織のもと)を、万遍なくまばらに緑の蛍光タンパク質でマークする方法を開発して、孵卵20時間以降に起きる脳の大まかな構造が作られる時期の細胞の変化を追跡しました。孵卵後25〜40時間の発生の時期に起きるプロセスです。動画1は10分おきにそれを観察した元のデータ、動画2は、マークされた(緑色に光る)細胞の軌跡を描いたものです。胚の前側の広い領域から胚の中心線に向かって、エピブラストの細胞が集まり、早く集まれた細胞が脳になり、遅れて集まってきた細胞が、脳を覆う表皮に発生することがわかりました。これらの動画には示されていませんが、細胞が集まる部分のエピブラストはAME(将来、脊索前板と前部脊索になる)という組織に裏打ちさせています。このような解析と、遺伝子発現の解析から、胚の前半分のエピブラストは、孵卵20時間ではすでに、「脳か頭部外胚葉*かいずれかに発生する」ように発生能が限定されていて、AMEにどこまで近く集まれるかで、脳と頭部外胚葉のいずれに発生するかが決まるらしいと推定しました。(*頭部外胚葉は、脳を覆う一層の細胞で、主には頭部の表皮になりますが、目、鼻、耳などの感覚器の組織や口から喉にかけての皮の部分も生み出します――眼科と耳鼻咽喉科の守備範囲の組織だと考えていただければ間違いありません。)
(動画1)
(動画2)
(動画1)ニワトリ胚の脳をつくる細胞の移動
ニワトリ胚の上側の層であるエピブラスト細胞をまばらに緑の蛍光(緑色蛍光タンパク質)で標識してタイムラプス(コマ落とし)撮影し、標識細胞の移動を目で追えるようにした13時間の記録。画面の横幅は4.5 mm。周囲のぼんやりとした緑は、胚を取り出したときにくっついて来た卵黄からの蛍光。エピブラストの端の方から胚の中心線に向かってエピブラスト細胞が集まって胚を作り、画面右側に脳が発生している。(1時間の変化を1秒で表示)
(動画2)ニワトリ胚の脳を作る細胞の移動の軌跡
(動画1)の緑で光るエピブラスト細胞のデータを軌跡として示した。込み入った線のそれぞれが識別できるように、軌跡ごとに色を変え、水平の青の線が頭の中心線となるように画面を整えた。脳を作る細胞が、胚の前側(画面右側)の広い範囲から中央に向かって集まることが確認できる。このデータをさらに分析すると、それぞれの胚領域でのエピブラスト細胞の移動の方向や速度、それらの経時的な変化を知ることができる。(1時間の変化を1秒で表示)
【動画1, 2の出典】Yoshihi et al. (2022) Development, doi: 10.1242/dev.199999
このことを確かめるために胚の前半分の端の方に赤に光るノード(画面ではマゼンタで表示)を移植すると、ノードそのものではなく、ノードから伸び出したAMEに緑の細胞が集まって第二の脳を作り、この場合でも遅れて集まった細胞が表皮になることがわかりました(動画3)。AMEだけを移植しても全く同じことが起きるので(動画4)、ノードは、AMEの供給源として働いているだけで、ノード自体には、オーガナイザーとしての働きは全くないことも明らかになりました。
(動画3)
(動画4)
(動画3)ニワトリ胚の脳の発生:ノードから伸びるAMEの働き
宿主ニワトリ胚のエピブラスト細胞を(動画1)の方法でまばらに緑色蛍光で標識した後、赤い蛍光で標識した(画面マゼンタ色)別の胚のノードを、宿主胚のノードの横の方に移植した。移植ノードには緑色の標識細胞が集まることはないが、ノードから画面右下方向に細長い組織(AME)が伸びだすと、マゼンタのAMEの周りに緑色のエピブラスト細胞が集まり(2〜5秒)、その集まったエピブラスト細胞が第2の脳を作る(6〜10秒)ことがわかる。(2時間半の変化を1秒で表示)
(動画4)ニワトリ胚の脳の発生:AMEにエピブラスト細胞が集合
(動画3)で示したノードの代わりに、伸び始めたばかりのAME(画面マゼンタ色)を移植すると、移植AMEは直ちに周りのエピブラスト細胞を集合させ、(動画3)と同様に第2の脳を作った。このことから、AMEへのエピブラスト細胞の集合が脳の発生の重要な1段階であることが確認された。(1時間の変化を1秒で表示)
【動画3, 4の出典】Yoshihi et al. (2022) Development, doi: 10.1242/dev.199999
さまざまな発生時期の異なったニワトリ胚の領域の働きを足し合わせると、両生類胚のオーガナイザーに近いものになる
鳥類や哺乳類の胚で原腸陥入(エピブラストの細胞が、その下層に落ち込んでゆく細胞移動)が起きているのは、原条とその端に生まれるノードですが、時間と場所で、それぞれ生み出すものが違います。ニワトリ胚で具体的に示すと次のようになります。
1. 孵卵14−17時間の原条:骨格や筋肉のもとになる中胚葉組織を生み出す。
2. 孵卵17時間あたりに原条の先端にできかけるノード:内胚葉組織(消化管の前駆体)を生み出す。
3. 孵卵20−27時間にノードから前方に伸びだすAME:エピブラスト細胞を集合させて、脳を中心とした頭部を生み出す。
4. 孵卵25−30時間のノード:後側の脊索を生み出す。
これら、時間も場所もバラバラの、エピブラストの下層への落ち込みに関わる胚領域の作用を足すとやっと、両生類でいわれる「オーガナイザー」に近いものになります。言い換えれば、ニワトリ胚には(そして鳥類胚、哺乳類胚一般についても)「オーガナイザー」というべき単一の組織は存在しないのです。さらに見方を変えれば、両生類の発生は特殊な組織構成のもとで進行するために、上に述べた1〜4の異なった組織とそれらの働きが、狭い領域と時間帯に凝縮されているために「オーガナイザー」と表現したくなるような胚領域が見出されるのではないかと考えられます。この考察は、私たちの実験の結果からもたらされたものですが、他の研究者も同じ結論に達しています。マウス胚と両生類胚を中心とした、現代的な(静かな革命のもとにある)研究結果を詳細に比較検討したAlfonso Martinez Arias(マウス胚の研究者)とBen Steventon(両生類胚の研究者)[6] は、「両生類のオーガナイザーが、異なった特異的な作用を持つ要素が偶然に集まってできたものであるのに対して、オーガナイザーを構成するそれらの要素は、他の(脊椎動物)種では、時間的にも空間的にも分散された状態で存在している。」と結論しています。両生類胚についても、個々の細胞の時々刻々の変化を追跡した解析を行えば、他の脊椎動物種との共通性が確認され、実験発生学に基づいた「オーガナイザー」説から抜け出すことができるのではないかと期待しています。
引用文献と原文
[1] Hans Spemann (1938) Embryonic Development and Induction.“I wish to emphasize that I have never constructed ‘an organizer theory.’ I coined the term ‘organizer’ to describe some new and remarkable facts when I ran across during my experiments.”
[2] Johannes Holtfreter and Victor Hamburger (1955). Analysis of Development Chapter 1. “Spemann (1921) introduced the term “organizer” for the upper blastoporal lip. The choice of this term, which we, like Spemann (1938) himself, shall use only in a provisional sense, was based on the discovery that the tendency of the graft was combined with the induction of the host tissue to form an integrated whole.”
[3] Johannes Holtfreter (1985) Reminiscences on the Life and Work of Johannes Holtfreter. Plenum Press. ”the organizer tissues do not actually organize the cell material whose new trend of differentiation they have induced. Rather the induced cells organize themselves into complex organs. Therefore, in later discussions of this issue, I went as far as to declare the term “organizer” to be a misnomer.”
[4] Victor Hamburger (1988) The Heritage of Experimental Embryology: Hans Spemann and the Organizer.Oxford University Press.
[5] Koya Yoshihi, Kagayaki Kato, Hideaki Iida, Machiko Teramoto, Akihito Kawamura, Yusaku Watanabe, Mitsuo Nunome, Mikiharu Nakano, Yoichi Matsuda, Yuki Sato, Hidenobu Mizuno, Takuji Iwasato, Yasuo Ishii, Hisato Kondoh (2022). Live imaging of avian epiblast and anterior mesendoderm grafting reveals the complexity of cell dynamics during early brain development. Development, doi: 10.1242/dev.199999.
[6] Alfonso Martinez Arias and Ben Steventon (2018) On the nature and function of organizers. Development 145, dev159525. “Whereas the amphibian organizer is a contingent collection of elements, each performing a specific function, the elements of organizers in other species are dispersed in time and space.”
バックナンバー
VOL.4 胚のいつ、どこで、何を発生させるのかには欠かせない抑制機構
VOL.8 動物の形の多様性という名の変奏曲集I 目と顔と手