

VOL.3 「細胞分化」の意味の変遷
近藤寿人(JT生命誌研究館 顧問・表現ディレクター)
発生生物学に関する解説や教科書を見ると、「細胞分化」という表現に沢山でくわします。この連載のVOL.1でお話しした「実験発生学」に基づいてさまざまな現象が解釈された結果、学術用語として細胞分化という表現が生まれました。「実験発生学」の欠点も「細胞分化」の考え方に反映されているので、(1)この表現が、どのようないきさつのもとで使われてきたのか、(2)この表現は現代の眼からはどのように再解釈する(正しい意味を与える)のが良いか、について検討していきましょう。
「分化」というのは、“differentiation”という外語に対してつけられた和語です。そのもとである“differentiate”というのは、違いを(目で見たり感じたりして)区別するという意味です。受精卵から胚、そして成体へと発生が進むと、その全体の姿だけでなく、それを構成する組織も目に見えてどんどん変化してゆきますから、「組織はどんどん分化(differentiation)を続けてゆく」というのは妥当であり、当初はそのような意味でした。
「分化(differentiation)」が、細胞集団である組織ではなく細胞に対して積極的に使われるようになったのは、細胞培養技術の発達によるものです。いくつかの種類の細胞については培養皿の上で単独で培養できるようになり、そのことによって、一つの細胞を上手く培養するとどのような細胞が生まれてくるかを追跡できるようになりました。細胞培養を用いた研究の初期の成功例として、筋芽細胞の培養と、血球系の細胞の培養が挙げられます。
筋芽細胞のクローン培養が扉を開いた現代的な細胞分化研究
細胞培養の技術が試行錯誤の最中であった1960年代に、金字塔ともいうべき研究を行ったIrwin R. Konigsbergの研究(1963)[1]を紹介しましょう。胚から取り出した1個の細胞がどのように発生してゆくかを調べる「クローン培養」の方法を編み出して研究に用いたのです。
発生途上の筋組織(たとえば孵卵10日のニワトリ胚の後肢の筋組織)には、たくさんの筋芽細胞(骨格筋のもとになる単核の紡錘状の細胞)とそれ以外の結合組織の細胞(線維芽細胞)とが含まれています。この筋組織を、タンパク質分解酵素トリプシンを用いてバラバラの細胞にまで解離します。(細胞自体は傷つけずに、細胞表面の細胞接着タンパク質だけを分解する程度の穏やかな処理を行います。)そして、1個1個の細胞が十分に離れる(シャーレあたり200個ぐらいの)密度で撒き、個々の細胞がシャーレの底にくっついたら、その位置をシャーレの裏からマークして、その変化を毎日写真に撮って追跡したのです。1細胞からの増殖によってできた細胞集団を、細胞クローンと呼びます。筋芽細胞についての、細胞クローンの日を追った変化の一例を、彼の論文の写真から描きなおした図1に示します。
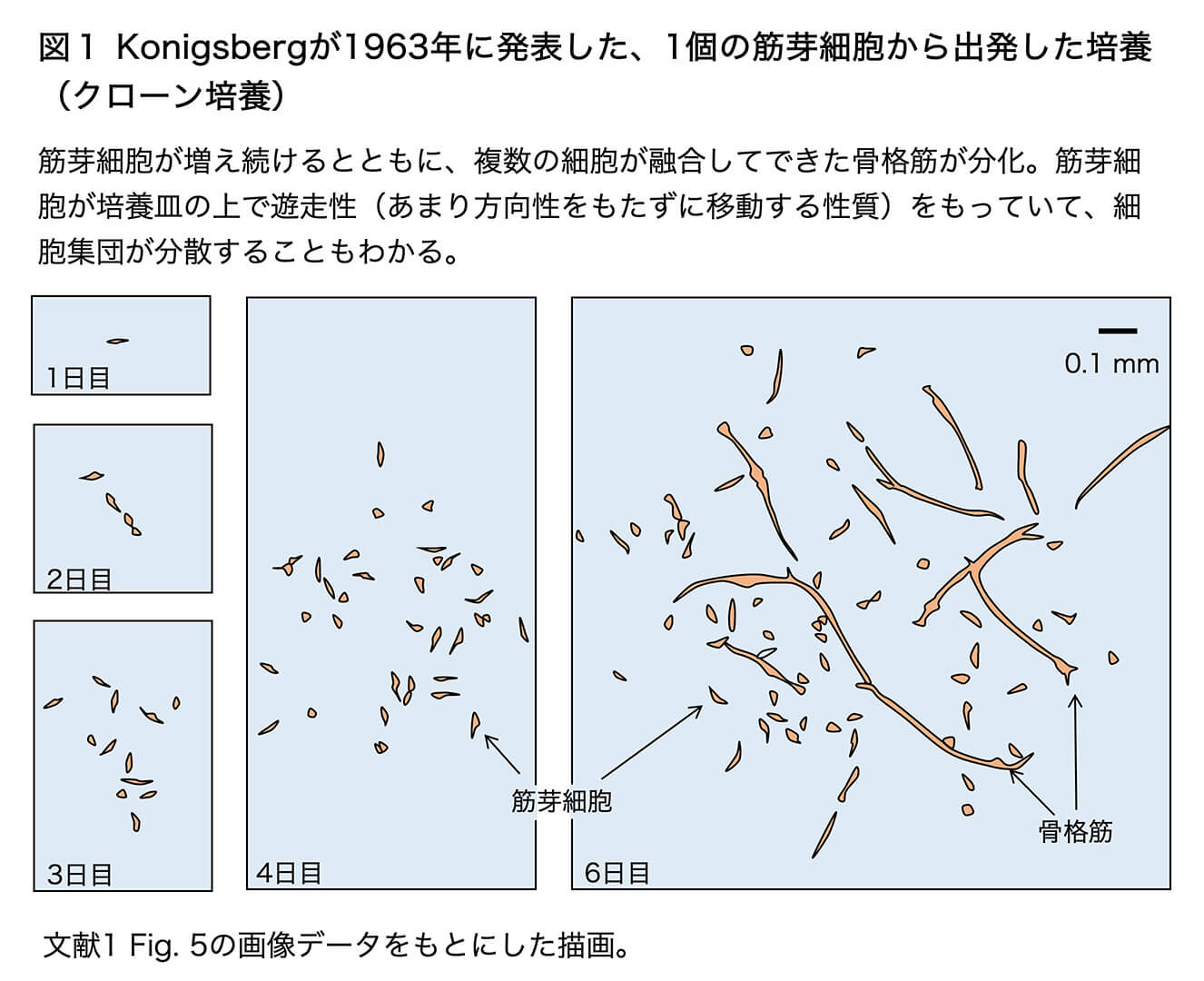
筋芽細胞は、培養4日までは、単核の紡錘形の細胞として1日に約4倍に数を増やすといったスピードで増えていきますが、5〜6日経つとそれらの一部が融合して、細長い多核の骨格筋繊維細胞に分化しました。骨格筋繊維の中の核はもはや分裂しません。培養の中では、隙間がある限り(永遠に近く)筋芽細胞が増え続けながらその一部が骨格筋繊維に融合して行って、巨大な骨格筋繊維の網目を作りますが、その隙間では必ず筋芽細胞が増えているという状態が続きます。筋芽細胞は、培養を続けても安定に「骨格筋繊維」になる(分化する)性質を失うことはありませんでした。一方で、筋芽細胞から繊維芽細胞が生まれたり、線維芽細胞から筋芽細胞が生まれたりすることも決してありませんでした。
「細胞分化」に関する教条的な理解の時代
培養された細胞の「分化」のいくつかの例から、たとえば筋芽細胞は骨格筋にしか分化しないので、骨格筋分化に「決定」されており、その「決定」状態に従って細胞は「分化」するのだーーといった考え方が広まり、どの細胞分化の場合にもそうだろうという考え方と、(VOL.2でお話しした)「特定の体細胞を生み出すには、特定の細胞系譜を辿らなければならない」という実は根拠のない発想とが結びついて、『細胞分化の「決定」がエピジェネティックランドスケープでの細胞系譜の分岐に対応する』などという考えまでもが一人歩きし始めました(Konigsbergには責任ありません)。その後、「決定」といってしまうと、次に述べる幹細胞系列の分岐の表現には相応しくないといったことなどから、「決定」に代わって「特異化(specification)」といった表現が用いられるようになりました。そこにも「細胞系譜」の重視が見られますが、その問題点はVOL.2でお話ししました。
ここで「細胞分化」に対する古典的な考え方を反映した用語についてお話ししておきます。
1. 「分化転換」。「特異化――>分化」と言う見方からは、VOL.2で取り上げた「網膜から水晶体」への発生などはうまく説明できないので、そのような例外的な現象は総じて「分化転換 (transdifferentiation)」と呼ばれました。しかし、実際には発現される転写因子の一部が共通した、細胞状態に関連性のある細胞への変化なので、私は「分化遷移」と言う表現を使いました[動画「生命(いのち)が形を作るとき」22分以降をご覧ください]。
2. 「脱分化」。これは主に2つの文脈で使われます。
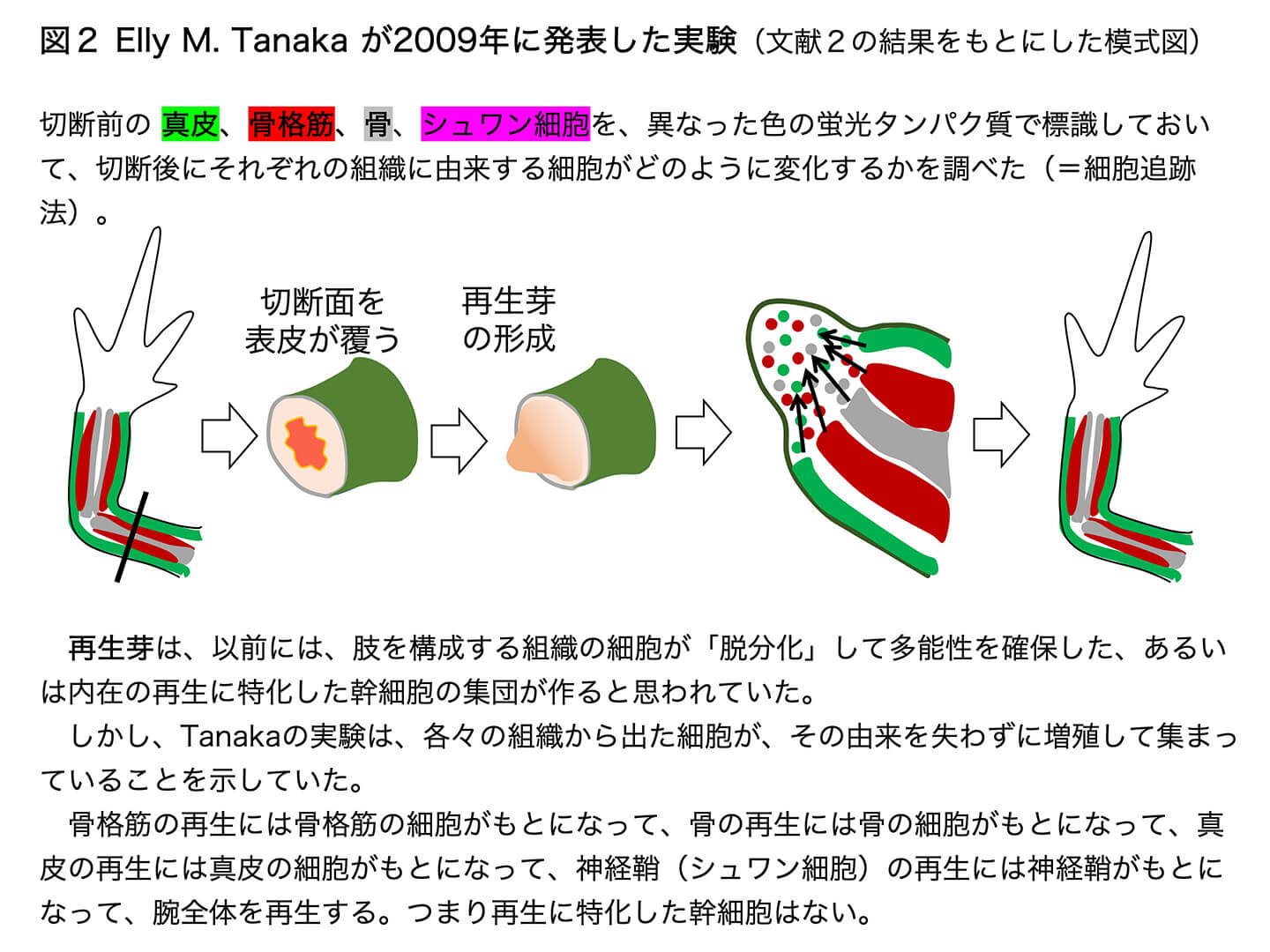
(1)発生の早い段階の、多分化能をもった細胞状態に戻ることが空想された場合。図2は、イモリの肢の切断後の再生の過程を示しています。切断した面が皮膚で覆われると、その皮膚の下に増殖が盛んな細胞が集まってきます。これらの細胞は、古典的な染色法で観察すると均一に見え、形の特徴もないことから、『ひとたび骨格筋や骨に分化した細胞が「脱分化」した結果できる、発生段階の初期に近い多能性細胞であり、肢の再生のための幹細胞として働く』と空想され、その考えがなぜか定着してしまいました。しかし、Elly Tanakaは2009年に、肢を切断する以前の段階の真皮、骨格筋、骨、シュワン細胞などを、異なった色の蛍光タンパク質で標識しておいて、切断後にそれぞれの組織に由来する細胞がどのように変化するかを調べる画期的な実験を行いました[2](図2)。その結果、骨格筋の再生はもとの骨格筋だった細胞がもとになって、骨の再生には以前に骨であった細胞がもとになって、真皮の再生には真皮であった細胞がもとになって、神経鞘(シュワン細胞)の再生には、以前に神経鞘であった細胞がもとになって、腕全体を再生することがわかりました。再生芽の中の細胞は、確かに骨格筋や骨の細胞に特徴的な形を示してはいませんが、胚発生の早い段階に戻っているわけではありません。再生という状況に備えて形には特徴を表していませんが、骨格筋や骨になる性質は安定して維持しているので、古典的な意味での「脱分化」は起きていません。
[動画「再生力のチャンピオン イモリとプラナリアのワザ比べ」の、22分から26分あたりも、あわせてご覧ください。]
(2)これも組織再生の時にしばしば見られることですが:(分化)状態Aの細胞からBの状態の細胞が作られる場合には、A細胞が「脱分化」して、A細胞とB細胞の細胞系譜の分岐点の状態にまで遡ってからB細胞に分化するという考え方。これは、特定の細胞種を生むためには特定の細胞系譜を辿らなければならないという、過去の教条的な考え方を反映するもので、事実に基づいたものではありません。多くの場合、AからBの直接の変化(例:虹彩から水晶体の再生)、あるいは細胞系譜の分岐点とは違う中間状態を経由するというのが本当です。後半にお話しする「1細胞トランスクリプトーム解析」の現状をご覧いただければ、細胞系譜は深い意味はもたないことを納得していただけるでしょう。
ちなみに皆さんは、上に出てきた「筋芽細胞」のほか、骨芽細胞、赤芽球(血球の細胞は「球」と呼ぶことが多い)、線維芽細胞など「芽」を含んだ細胞の名前に出くわすことが多いと思います。「芽細胞」は、増殖可能状態にあり(常に増殖していなくても良い)、特定の細胞種だけ(例えば骨格筋)だけを生み出す細胞を表しています。よく皆さんが目にする「線維芽細胞」は、繊維細胞(タイプIコラーゲンを産生する細胞)のもとになる増殖状態の細胞です。
一方で、次に登場する「幹細胞」は、増殖可能状態にあるとともに、それから最終的には新たに2種類以上の細胞種を生み出す細胞です。神経幹細胞からは、ニューロン(神経細胞)とグリア細胞の両方が生まれます。
造血系の研究が細胞分化の理解に大きな貢献をした
細胞培養が発生過程の研究に大きな貢献をした例としては、血球系の発生(造血系)の研究を欠かすことはできません。造血系の細胞の培養には、免疫系の発生の解明、白血病の研究などを目的として国際的にも最先端の多くの研究室が参加しました。私が駆け出しだった1980年代初頭の、発生研究の国際集会には必ずといっても良いほど造血系の発生の演題が招待されていました。
造血系の細胞を研究する利点はたくさんあります。(1)造血組織(骨髄など)から細胞集団をとってきた上で、その中から特定の発生段階にある細胞を、細胞表面にあるタンパク質(=表面抗原)の違いをもとにして、比較的簡単に分別して集めてくることができること。(2)その分別された細胞集団を培養すると、それが増殖性の細胞なのか、どのような細胞を生む(分化する)のかなどを知ることができること。(3)造血組織から分別された細胞であっても、培養された造血系の細胞であっても、それらの細胞をX線照射マウス(造血幹細胞を失ったマウス)などに血管を介して移植して、どこまで造血系を回復させることができるかを確認し、それから遡って、それらの細胞の発生段階の中での立ち位置を推定することができること、などです。
その造血組織の発生の現時点での理解の状況を見てみましょう。ヒト(あるいはマウス)の造血組織(骨髄など)、臍帯血、末梢血のいずれをとってみても、血球系の細胞の全ては、造血幹細胞(Hematopoietic stem cell, HSC)を起源として作られます。HSCから出発して、どのような中間段階の細胞を経て、各々の血球(血液細胞)が生まれてくるのか(分化するのか)を整理した、現時点での概略図を図3に示します(文献3に基づく)。これは定説というよりも多数意見といった程度のものです。いずれにせよ、この図の右端や左端に置いたさまざまな血球は、もはや分裂して増殖することはなく、寿命があります。血球の寿命は長いもので赤血球の4ヶ月ほど、寿命が短い白血球では数時間から数日です。これらの血球は常に新しいものに置き換えられています。
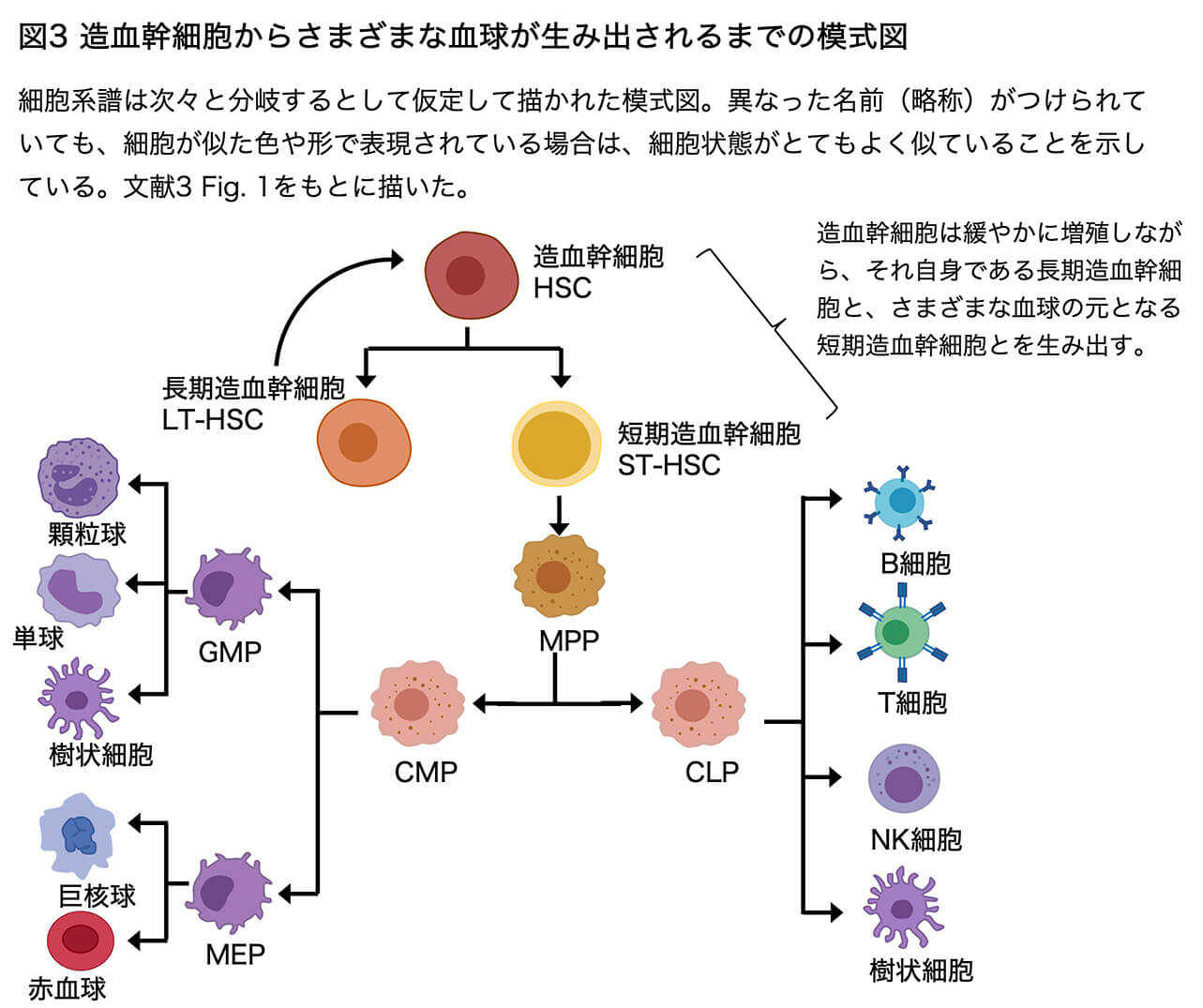
図3のように整理してしまうと、「やはり細胞系譜は重要ではないか」という印象をもたれるかもしれません。しかし、ある段階から次の段階への変化、あるいは変化した後の細胞種の間の区別は、そんなに歯切れの良いものではないのです。図3はあくまでも、「新しい細胞種が生まれる時には、細胞系譜が分岐してゆく」という仮定のもとに描かれたものに過ぎません。一応異なった名前(略称)がつけられていても、細胞が似た色や形で表現されている場合は、細胞状態がとてもよく似ていることを表しています。それらの間の区別は微妙なものです。またこの図において、同じ樹状細胞(dendritic cells)が、右側と左側の分枝の細胞系譜のいずれによっても生み出されるように表されている(特定の細胞系譜には依存しないようだ)ことにも注目しておいてください。
1細胞トランスクリプトーム解析がもたらした、細胞分化の理解の革命
最近の10年間に起こった、生命科学分野での目覚ましい技術革新は、1個1個の細胞の中で合成されているmRNA(転写産物)を、塩基配列解析によって網羅的に調べることができるようになったことです。この技術は、1細胞トランスクリプトーム解析(トランスクリプト=転写産物 + オーム=まるごと)、あるいは技術面を強調してsingle cell RNA-seq analysis などと呼ばれます。これを、数千から数万の細胞について行います。(現状では、細胞を1個1個にバラバラにして解析するので、もとの細胞集団のどこにいた細胞なのか?という情報は失われますが、その情報を補う工夫はあります。以下で例としてあげる造血系では、もともと細胞はバラバラです。)その次の作業として、発現されている遺伝子の種類とそれぞれの発現量から見て、似た状態の細胞は近接した点として、また状態が隔たった細胞は離れて位置した点として、多数の細胞の関係を2次元上で表示します。(このために、いろいろな計算方法が工夫されていますが、次に示す図4では、SPRINGという方法を用いた例を示しています。)そして、その2次元上の細胞状態を表す点の分布から、どの状態からどの状態に細胞状態が遷移するのかを推定することができます。
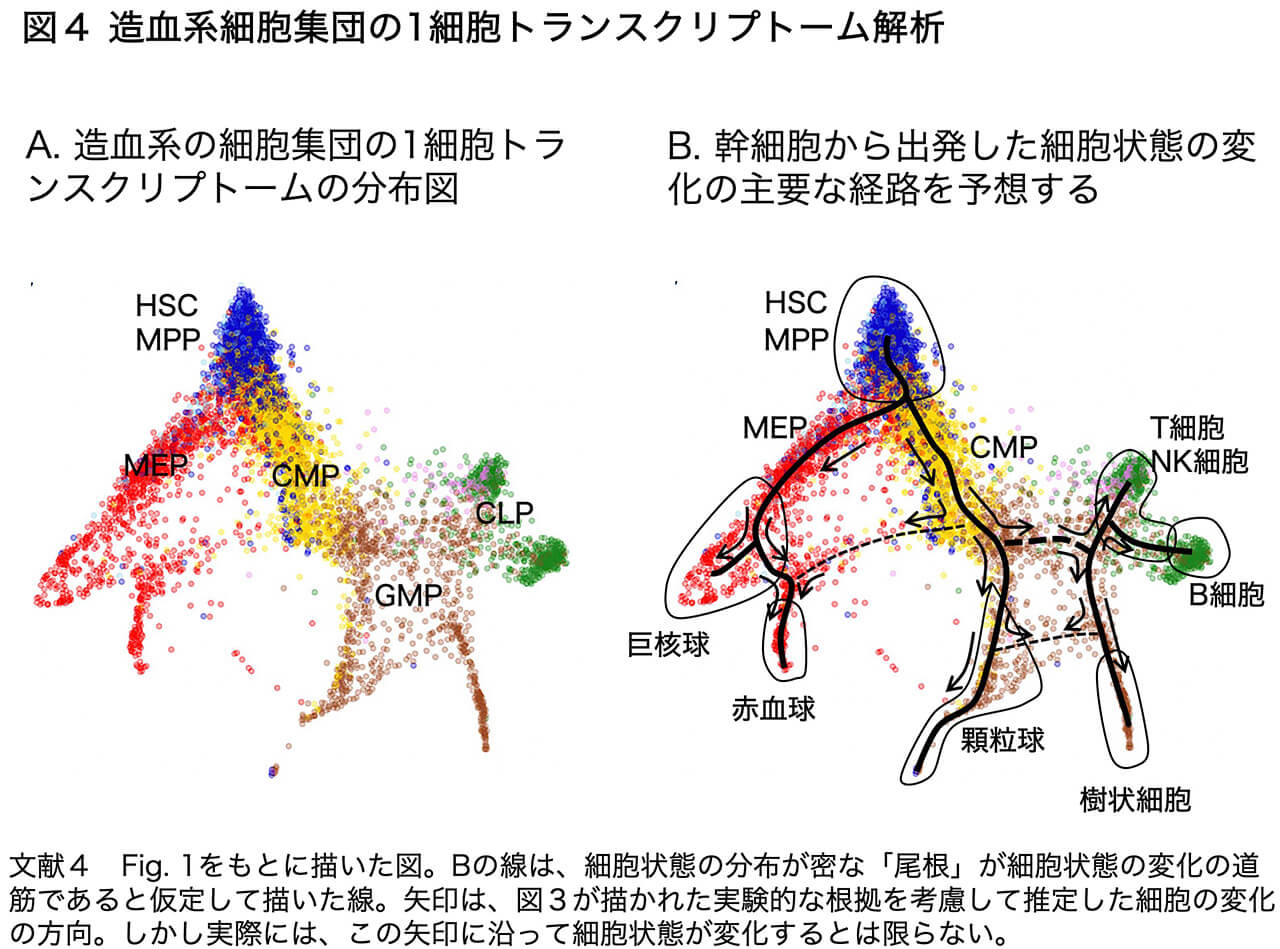
図4は、ヒトの造血組織である骨髄の細胞について、単一細胞トランスクリプトーム解析を行った結果を示しています(文献4 Fig. 1を改変)。図3で示した細胞種は網羅されています。左のAのパネルは細胞種についての解析の前の状態で、造血幹細胞(HSC)、MPP、CMP、CLP、GMP、MEP細胞の表面抗原をもつ細胞がどのように分布しているかを色分けして示しています。まず造血幹細胞とMPPの分布が重なったことから、この2つの細胞種の間で、細胞状態には大きな違いがないことがわかります。またこのAのデータから、「MPPからまずCMPとCLPという2つの前駆体が分かれる」という図3の整理は、現実を反映してはいないようだということがわかります。右のBのパネルは、Aのデータに対して、2つの作業を加えたものです。(1)「赤血球はヘモグロビンを産生している」といった基準から、どの範囲の細胞状態が「分化した」血球集団に対応しているのかを示しました。(2)細胞密度が高い「尾根」を辿ってゆくことによって、ある細胞状態から「主には」どのような中間段階を経て「分化」細胞(血球)に至るのかを推定しました。太い実線は、コンピュータープログラムでその峰を描かせたもの。それに、低い尾根を伝っての細胞状態の遷移の可能性を(他のデータに即して)、太い破線(論文の著者による)および細い破線(筆者による)で追加しました。これらの尾根の配置を見ると、「幹細胞から細胞系譜が分岐しながら進み、最終的な(分化)状態に至る」といった古典的な世界観とは大きく外れて、「幹細胞から最終的な(分化)状態に至るには、複数の道筋がある」ことが予想されます。さらに、これらの「尾根」から離れた位置にも、相当数の細胞状態が分布していることから、「尾根」を細胞状態の遷移の幹線道路であるとみなすと、さまざまな迂回路や短絡路もあることが示唆されます。またBのパネルでのMEPやCMPの細胞状態の分布が広いことから、これらの中間段階の細胞群は一様ではなくて、徐々に次の細胞状態への変化を進めているさまざまな段階の細胞の総称というべきものであることもわかります。
図4から、さまざまな実験データがどのように解釈されて図3が描かれたのかということは推定できますが、事実を表しているのは図4の方なのです。これからの時代では、「細胞分化」と呼ばれた発生のプロセスは、図3のような発想ではなく、図4が示す事実に基づいて理解しなければなりません。特に「細胞系譜が分岐していくことによって初めて、さまざまな形質(特徴や機能)をもった細胞タイプが生まれる」という従前の考え方は、真実を見誤らせます。
血球系の研究は、一生にわたって血球系を生み出し続ける造血幹細胞から、末梢の多様な血球に至るまでの全ての発生過程を全貌できるので、「細胞分化」の理解に革命的な変化を起こした、1細胞トランスクリプトーム解析の好例として取り上げました。血球以外の組織でも、1細胞トランスクリプトーム解析は同じ結論をもたらしつつあります。ある細胞状態の前の状態が何であったかを確認するために、文献2で行なったような細胞標識を行ったり、組織の中の位置に対応して発現が変化する遺伝子の発現と1細胞トランスクリプトームとを関連付けたりするなどの、さまざまな工夫がなされています。少し観点を変えれば、特定の細胞タイプを生まれさせるための細胞系譜は限定されたものではないということです。VOL.2の最後のところで、ES細胞を培養した場合に見られる発生の経路が、胚発生の場合と異なっていることが少なくないことをお話ししましたが、1細胞トランスクリプトーム解析と同じ結論に到達していたのです。
『個体を作る胚発生では、さまざまな細胞タイプや組織の発生について、そのタイミングや空間配置を厳密に調節して、組織の複合体としての「機能する臓器」を作り、そしてそれらの間をつなぐ脈管系や神経系を完成させなければなりません。』このように言うと、やはり細胞系譜の限定が重要であるように聞こえるかもしれません。確かに大まかなところだけを見れば、先に述べたような、細胞の変化の経路の「幹線道路」が目立ちますが、細部の仕上げや、損傷を受けた組織の再生過程では「迂回路」や「短絡路」を活用しているように見えます。再生過程で見られる「分化遷移」は、迂回路を使った発生過程なのです。
引用文献
[1] Konigsberg IR. Clonal analysis of myogenesis. Science. 1963 140(3573):1273-84. doi: 10.1126/science.140.3573.1273.
[2] Kragl M, Knapp D, Nacu E, Khattak S, Maden M, Epperlein HH, Tanaka EM. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature. 2009 460(7251):60-5. doi: 10.1038/nature08152.
[3] Mann Z, Sengar M, Verma YK, Rajalingam R, Raghav PK. Hematopoietic stem cell factors: Their functional role in self-renewal and clinical aspects. Front Cell Dev Biol. 2022 10:664261. doi: 10.3389/fcell.2022.664261.
[4] Pellin D, Loperfido M, Baricordi C, Wolock SL, Montepeloso A, Weinberg OK, Biffi A, Klein AM, Biasco L. A comprehensive single cell transcriptional landscape of human hematopoietic progenitors. Nat Commun. 2019 10(1):2395. doi: 10.1038/s41467-019-10291-0.
図の引用
サムネイルは、文献4 Fig. 1より
バックナンバー
VOL.4 胚のいつ、どこで、何を発生させるのかには欠かせない抑制機構
VOL.8 動物の形の多様性という名の変奏曲集I 目と顔と手