

VOL.6 肺の発生にまつわる、2つの話題
近藤寿人(JT生命誌研究館 顧問・表現ディレクター)
今回は、肺の発生にまつわる2つの話題についてお話しします。
最初の話題は、私たちが呼吸器として持っている肺が食道から分かれて生まれる過程についてですが、これまた以前にお話しした「抑制することが大切(VOL.4)」ということの再確認でもあり、同時に発生生物学でよく語られる「上皮―間充織間相互作用」(これまではほとんど、間充織からの作用ばかりが語られていながら「相互作用」と言われていた)の上皮からの作用の証明でもあります。
第2の話題は、魚類が持っていた肺と、私たち4足動物が持っていた肺の関係で、生命誌研究館のシンボル的な展示動物である肺魚も主役です。
肺と食道の関係:食道の上皮でSox2の発現を無くすとどうなるか?
上皮と間充織の関係
喉の奥から肛門までをつなぐ消化管の一番内側の管は、1層の内胚葉の細胞層(一層の隙間のない細胞層なので、上皮と言います)が閉じてできた管です。その周りに、主には中胚葉の側板と言われる細胞群から発生した細胞――それを一まとめにして間充織と呼んでいるーーがぐるっと巻きついて、食道、胃、腸といった消化管ができています。
どこからが内胚葉なのでしょうか?病院の診療科で「耳鼻咽喉科」というのがありますが、この診療科は「外胚葉」がもとになってできた器官を対象としており、それよりも奥に入ると内胚葉の世界と理解していただければ良いでしょう。
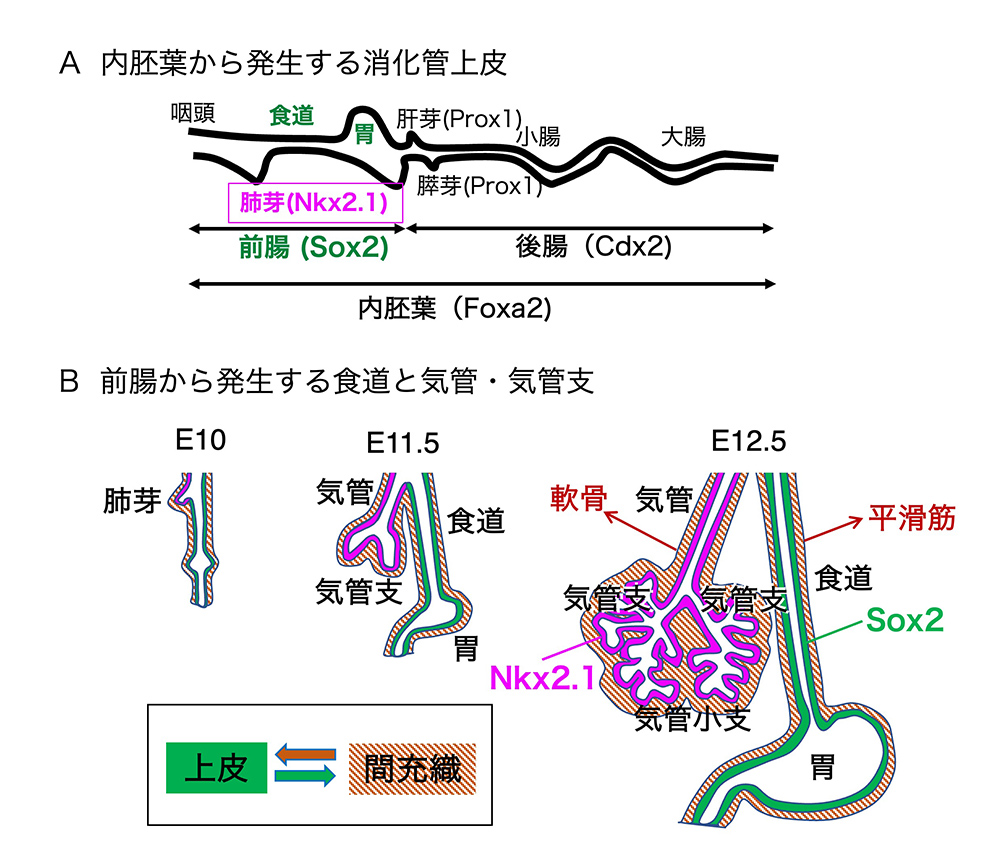
(図1)消化管の初期発生
胚の中で消化管を作る上皮の管ができた頃の、管の模式図を図1Aに示します。管は頭側から、食道、胃、十二指腸、小腸、大腸と続きます。Foxa2という転写因子が内胚葉由来の消化管上皮全体で発現されます。食道から胃までの「前腸」の上皮で転写因子Sox2が、十二指腸から大腸に至る「後腸」の上皮ではCdx2が発現されていて、消化管を二分する領域の上皮の性質を決めています。
食道の一部、十二指腸の一部に、肺芽、肝芽、膵芽という出っ張りができ、それらが肺、肝臓、膵臓へと発生してゆきますが、ここでは、肺芽から肺への発生を見てゆきましょう。
肺芽ではSox2に加えて、Nkx2.1という転写因子が発現され、Nkx2.1は、肺芽が気管に発生し、さらに気管から気管支、気管支から気管小枝と分岐が続く呼吸器系の上皮に発現し続けます。
間充織を含んだ、食道と呼吸器系全体の発生の様子を図1Bに示しました。Eのあとの数字は、マウス胚の受精後の日数で、ヒトの発生で言えば、妊娠1〜2ヶ月の時期にあたるとお考えください。食道を取り巻く間充織は後に、飲み込んだものを食道の喉側から胃側まで送り込むための平滑筋などになります。一方、気管・気管支の間充織は、のちに軟骨などになります。息を吸うときの陰圧に負けて気管や気管支がつぶれると呼吸できないので、気管・気管支の周りに洗濯機の排水ホースのように見える軟骨を発生させるのです。E12あたりの胚の間充織は、外観ではそのような差があるようには見えません。しかし、そこで発現している遺伝子を調べるとすでにはっきりした差があって、食道の周りでは平滑筋の発生が準備され、気管・気管支の周りでは軟骨の発生が準備されていることがわかります。つまり、Sox2だけを発現する食道の上皮の周りの間充織と、Nkx2.1を発現する呼吸器系の上皮の周りの間充織は(おそらく、上皮の違いの結果として)性質を変えているのではないかと思われます。
これまで、色々な臓器(必ず上皮と間充織からなる)は、上皮と間充織の相互作用によって発生すると言われながら、その実例として示されるものはほとんど例外なく「間充織が上皮の性質を決める」ケースであったために、「上皮は無垢であり、間充織の性質に従って変化する」といったイメージさえ蔓延している状況もありました。
私たちは、Sox2が食道と肺の発生にどのような役割を果たしているのか?という問題を解くために、「内胚葉だけでSox2を失わせたらどうなるか?」という実験を、マウスを用いて行ったのですが、その実験では同時に、「上皮の性質が間充織の性質を決めるケース」を具体的に示すことができました(文献1)。そのお話しをします。
マウス胚の内胚葉だけでSox2遺伝子を失わせる
ゲノム配列からSox2遺伝子を除くには、LoxPという配列(図2Aの黒三角)をSox2遺伝子の両側に挿入します(この状態のSox2遺伝子を、Floxed Sox2と呼びます。 Flanked by LoxP =“LoxPに挟まれた”の略です)。これにCre組み換え酵素が作用すると、Sox2を含む配列が環状DNAとなって、ゲノム配列から除かれてしまうのですが(図2A)、Cre酵素を「内胚葉だけで」働かせるために、次の工夫をします。(1)CreERT2という、4-OHタモキシフェンが結合すると初めて活性を持つCre酵素を用いる。(2)CreERT2を内胚葉で発現するFoxa2遺伝子と同時に発現するような遺伝子操作をする(この詳細は、今回は省略します)。Floxed Sox2をホモの状態でもち、Foxa2と同時にCreERT2を発現する胚を、図2B下部に示した雄雌の間の交配で準備しておき、Foxa2が初期胚での役割を終えて、新生の内胚葉での発現に移行したE7.5の発生ステージをねらって母親にタモキシフェンを経口投与します(念のためにE8.5でも)。すると、母親の肝臓で代謝されてできた4-OHタモキシフェンによってCreERT2が作用して、胚の内胚葉でSox2遺伝子がゲノムから切り離されて活性を失います。
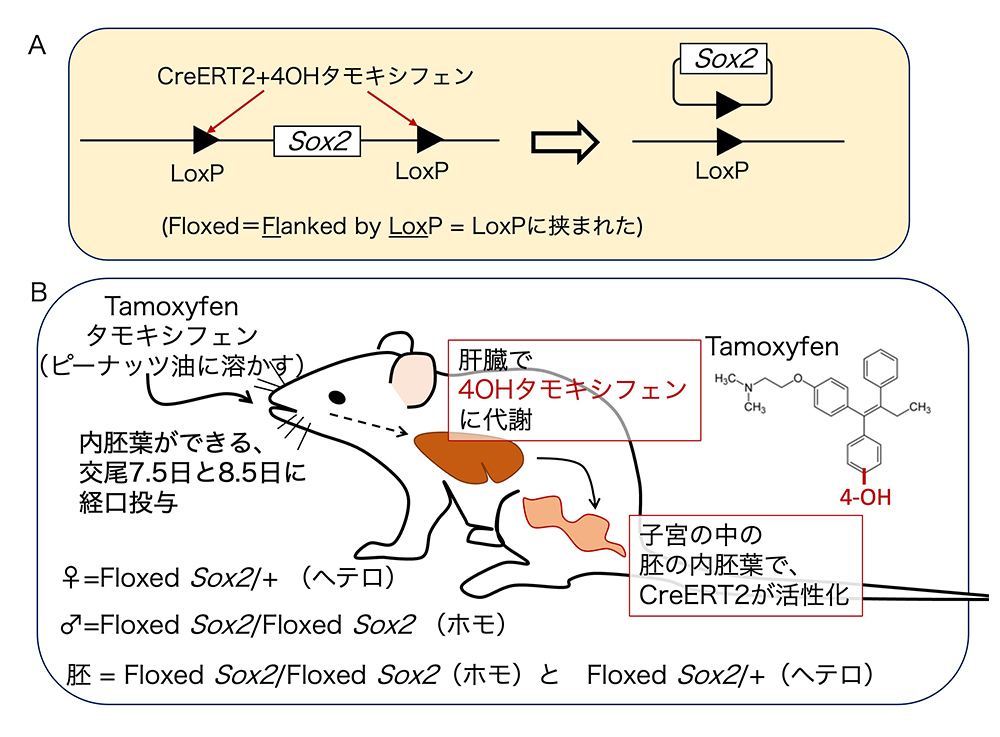
(図2)胚の内胚葉でSox2遺伝子を欠失させるための遺伝子操作。
A. Floxed状態の Sox2 遺伝子に、活性を持ったCre組換え酵素が働くと、遺伝子が環状DNAとして切り除かれる。
B. 妊娠雌マウスにタモキシフェンを与えてから、子宮の中の胚の内胚葉でCreERT2が4-OHタモキシフェンによって活性化されるまで。
内胚葉でSox2遺伝子を失った(∆Sox2)マウス胚の前腸の発生
上に述べた遺伝子操作の結果、内胚葉だけでSox2遺伝子を失った前腸はどのように発生したのでしょうか?マウス胚には、Fox2と同時にEGFPという緑の蛍光タンパク質を発現させるという、もう一つの遺伝子操作を行っていて、食道や気管などの内胚葉由来上皮が緑に光ります。
妊娠13.5日で取り出した胚の組織が図3左側に示されています。喉から胃までをつなぐ細い管が食道です。それとは別に、喉から気管が伸び、それがさらに気管支に分かれ、その先はさらに細かく分枝して、呼吸器特有の組織を作っています。
内胚葉でSox2遺伝子を失ったマウス胚(∆Sox2)の前腸を図3右側に示します。喉から出る上皮の管はたった一本で、少し小振りの胃につながっています。胃につながっているので、一見食道のようですが、驚くべきことに、その管から一対の気管支が伸びだし、その先は呼吸器特有の分枝が続いています。
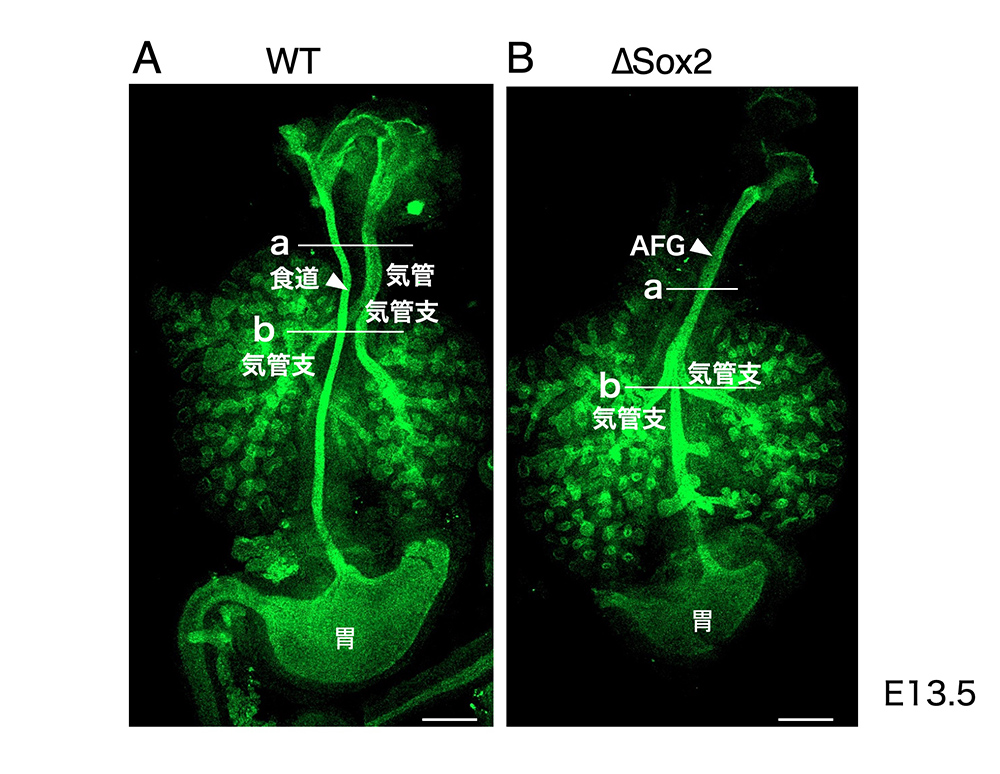
(図3)前腸上皮でのSox2の発現が失われたマウス胚では、食道と気管が分かれず、1本の咽頭と胃をつなぐ管(AFG)から1対の気管支が分枝する。(文献1 Figure 1Aを改変)。白線は500 μm.
では、この喉と胃をつなぐ前腸(AFG)上皮の管はどのような性質を持っているのでしょうか?それを調べるために、図3の(a)または(b)の場所の切片を作り、免疫染色で転写因子の発現を調べたり、in situ hybridizationで遺伝子のmRNAの発現を調べて、正常胚の場合と比べました(図4)。
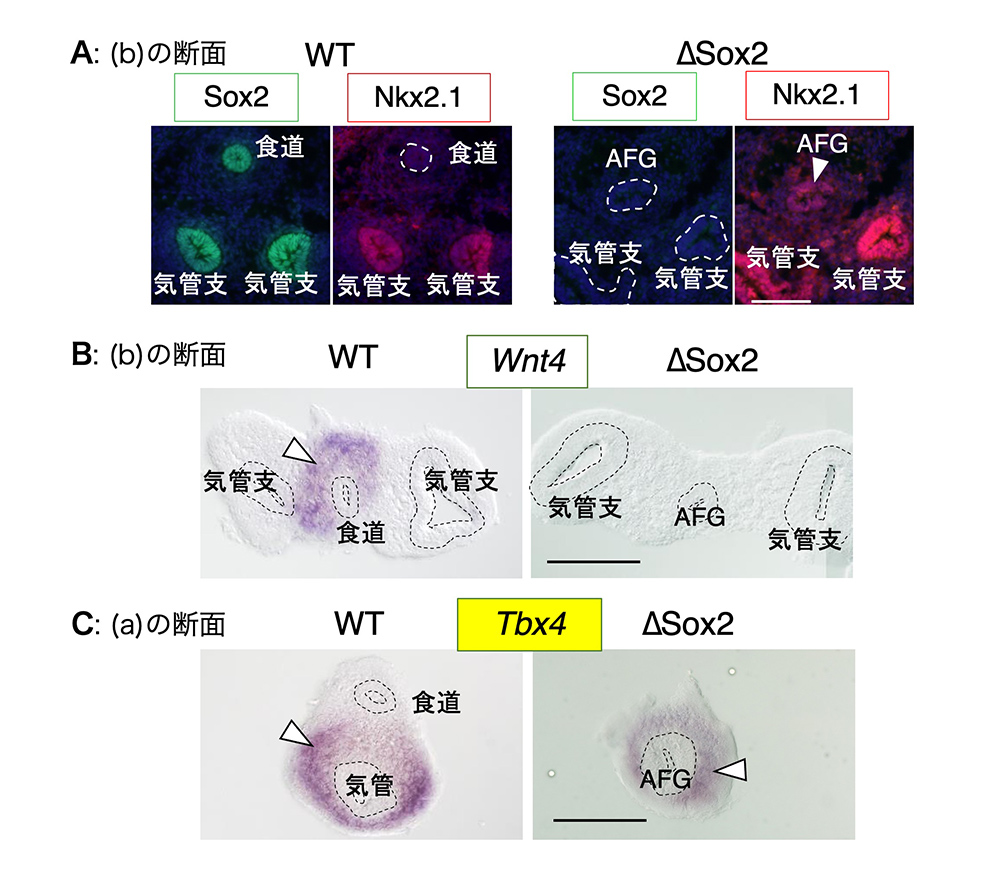
(図4) 前腸上皮(AGF)でのSox2の発現が失われると、食道になるはずだったAFGの上皮も周囲の間充織も食道の性質を失い、気管・気管支の性質を持つ。断面は、図3で示した位置(文献1 Figure 1BC, Figure 3BCを改変)。白線は100 μm.
正常胚の上皮はすべてSox2を発現し、呼吸器(気管支)の上皮だけがNkx2.1を発現しています(図4A)。一方、∆Sox2の胚では、全ての上皮がSox2の発現を失うとともに、全ての上皮の管が、呼吸器固有のNkx2.1を発現していました。このことから、内胚葉でSox2遺伝子を失った∆Sox2の胚の、喉と胃を結ぶ管(AFG)は、(食道の代わりに)気管の性質を持ってしまったことがわかりました。
では、その管(AFG)の周りを取り囲む間充織の性質はどうでしょうか?正常胚の染色(図4BC左側)で示すように、食道の間充織はWnt4遺伝子、呼吸器の間充織はTbx4を発現するのが特徴です。ところが、∆Sox2のAFGの管の周り間充織にはWnt4遺伝子の発現がない一方、Tbx4が発現されていました(図4BC右側)。つまり、喉と胃を結ぶAFGがSox2の発現を失ったために上皮としての性質が食道から気管に変化しただけでなく、それを取り巻く間充織も、気管の間充織としての性質を持ってしまったことが分かりました。
今回の記事の最初の方でのべた、上皮―間充織の相互作用のうち、「上皮から間充織へ」向けての作用の例を示すことができたのでした。
Wntシグナルが間充織に作用すると、Tbx4発現を引き起こし、それが組織の呼吸器系としての発生を促す
食道の上皮で発現されているSox2は何をしているのかについて、いくつかの研究を総合すると(文献1〜3)、次のようになります。Sox2は、食道上皮に食道としての性質を与えるとともに、Sfrp2という、Wntシグナルを強力に抑制する分泌タンパク質を合成します。
実は、食道の間充織も、気管・気管支の間充織も、Wntタンパク質を分泌しているのですが、食道の間充織では、Sfrp2の作用でWntシグナルが抑制され、食道の間充織としての性質を示します。
気管・気管支の間充織でTbx4遺伝子が発現されますが、Tbx4は間充織のWntシグナルによって発現される遺伝子なのです。Tbx4は転写因子で、Fgf10などの分泌タンパク質の遺伝子を活性化し、それらの分泌タンパク質が上皮に作用して、呼吸器の上皮として発生させます。
食道になるべき喉と胃をつなぐ上皮の管でSox2の働きがなくなると、食道上皮が気管上皮へと性質を変えるとともにSfrp2の分泌もなくなるために、間充織自身のWntシグナルが間充織でのTbx4の発現を引き起こし、全体としての呼吸器系組織としての発生(気管支の分岐等)に至るというわけです。
(注意深くデータをご覧になった方は、図3で示された気管や気管支でNkx2.1とともに発現されるSox2の働きはどうなっているのか?という疑問を抱かれるかも知れません。気管が食道と別れた後の呼吸器系上皮のSox2の発現には、これといった機能がなく、肺胞ができて呼吸器としての機能が成熟する段階でもう一度働くSox2のための待機発現ではないかと考えられます)。
次の話題の基礎として、ここでは『呼吸器を作る間充織では、WntシグナルによってTbx4が発現される』ことを、確認してください。
魚の肺はどうなっているのか?
ポリプテルスの肺
「魚の肺」と聞くと、おや?と思われる方が多いのではないでしょうか?魚は鰓(えら)呼吸で、あるとすれば鰾(浮き袋)くらいのものだと。確かに、鮮魚店の店先を占める「硬骨魚」はそうかも知れませんが、図5に示す系統樹と肺の図の対応でご理解いただけるように、魚類の多くの系統は「肺」と言われる、食道とつながった器官を持っています。「鰓呼吸だけでは、環境の水の中の(溶存)酸素が低下すると生命の危機に瀕するので、口から取り込んだ空気の酸素も利用する」という説を読んだことがあります。ガソリンエンジンとバッテリーの電気を共用するハイブリッド車のようなイメージです。
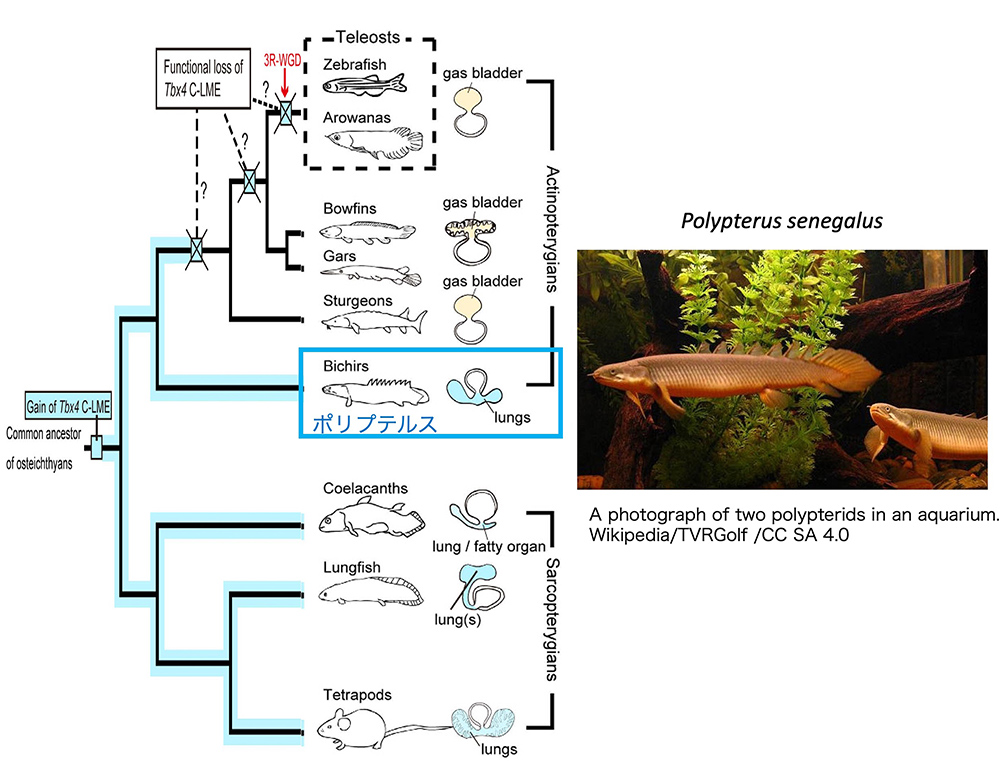
(図5) 魚類の系統樹と、魚類が持つ「肺」の模式図。
左側は、文献4 Figure 5を改変。右側は研究の対象となった、アフリカのポリプテルスPolypterus senegalusの姿。
ここでは、魚類の中でも大きな「肺」を持ったポリプテルス(古代魚とも言われる)の肺が、本当に私たち4足動物(陸に上がった脊椎動物)の肺と相同なのか?ということを調べた、慈恵医科大学の岡部正隆教授のグループの研究を紹介しましょう(文献4)。
ポリプテルスの胚発生で、丁度「肺芽」が食道から分かれる時期を選んで、間充織にTbx4, Tbx5の遺伝子が発現されるかどうか (mRNAが作られているか) が調べられました。Tbx5はTbx4とほとんど同じ発現制御をうけ、転写因子としても同等の機能を持ちます。すると、ポリプテルスの胚でも(マウスの場合と同様)、食道を取り囲む間充織には、Tbx4、Tbx5の発現はないが、それから別れてできた肺芽の間充織ではTbx4、Tbx5が発現されていました(図6)。
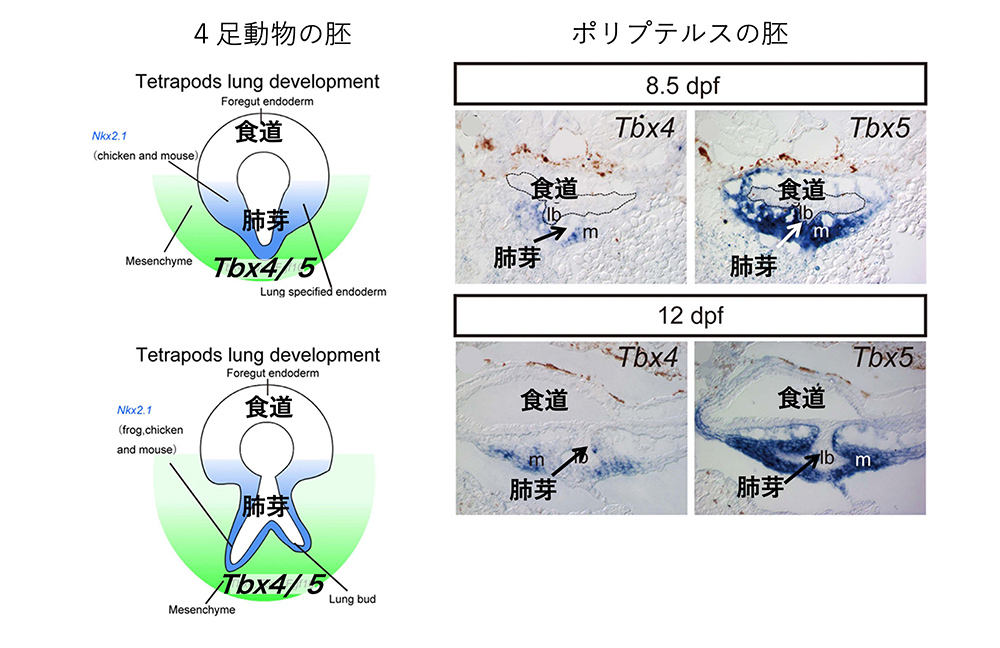
(図6)4足動物でもポリプテルスでも、胚の食道(前腸)上皮を取り囲む間充織ではTbx4は発現されないが、『肺芽』の間充織では発現される。文献4 Figure 2を改変。
さらに、魚類肺の間充織でのTbx4の発現制御が調べられました。マウスの研究から、Tbx4遺伝子のイントロン配列の中に、肺の間充織でTbx4を発現させるためのエンハンサー(lung mesenchyme enhancer, LME)があることが知られていました(文献4)。そのLMEとほとんどの塩基配列が、ヒト、ニワトリ、カエルのみならず、シーラカンスやポリプテルスのTbx4遺伝子のイントロンにも見つかりました。それらの配列がエンハンサー活性を持つかどうかを調べるために、エンハンサー活性があればEGFPを発現し、緑色に光らせるベクターtkEGFP(文献5)につないでニワトリ胚に導入すると、肺の間充織で緑に光り(図7B 緑色蛍光)、肺間充織でエンハンサー活性を示すことが確認されました。これらのエンハンサーには、Lef/Tcf1転写因子が結合する特徴的な塩基配列が同じ場所に現れます(図7C)。この配列は、LMEエンハンサーがWntシグナルによって活性化されることを示しており、前節で述べた、『呼吸器を作る間充織では、WntシグナルによってTbx4が発現される』ことは、ポリプテルスの肺の発生でも起きているのです。
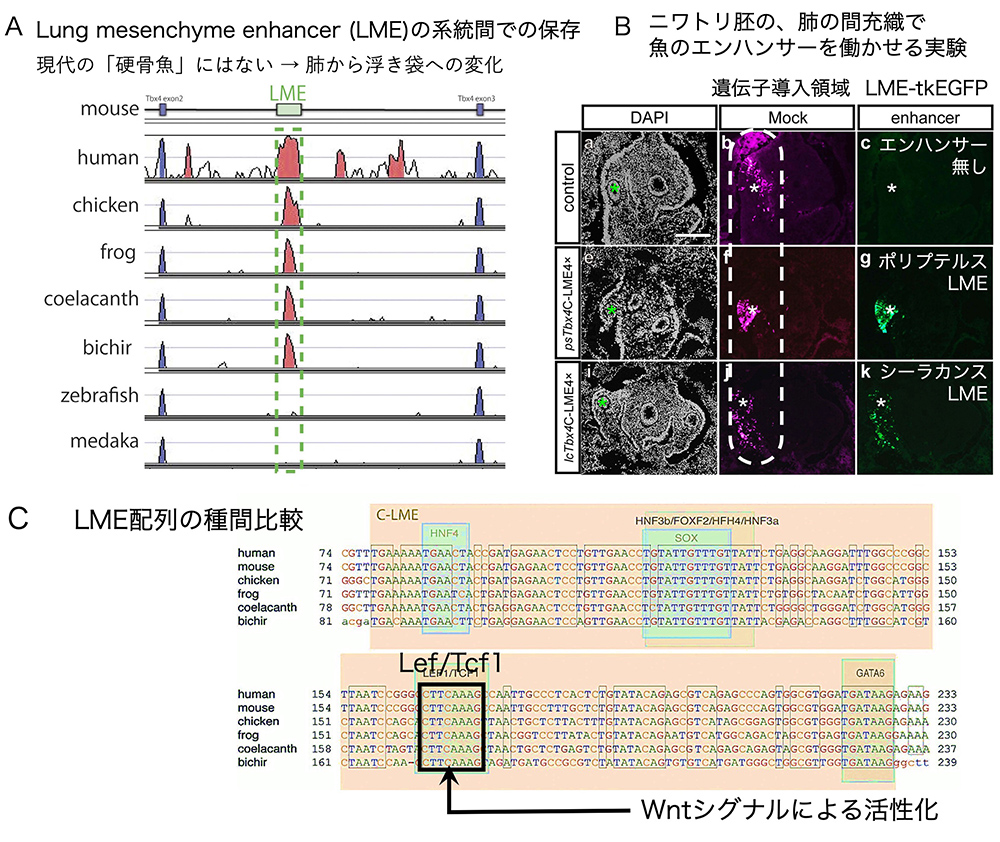
(図7)Tbx4遺伝子を胚の間充織で、Wntシグナルによって活性化するエンハンサーLMEは、シーラカンス・ポリプテルスなどの「古代魚」のTbx4遺伝子は持っている。(文献4 Figures 3, 4を改変)
興味深いことに、硬骨魚(ゼブラフィッシュ 、メダカ)のゲノムには、そのエンハンサーは見つかりませんでした(図7A)。硬骨魚では、浮き袋が肺のように食道から分かれてできるのですが、やがて食道からは離れて、空気を内蔵した袋状の器官として独立します。100年以上前から、浮き袋と肺が相同器官ではないかと考えられていましたが、当時は、未発達な浮き袋から肺が発達してできたという考えが主流でした。実際には、硬骨魚では、肺の間充織で発現されるべきTbx4が発現されないために、浮き袋になってしまったのです。実際、マウス胚の呼吸器の間充織でTbx4、Tbx5の発現が下がると、分枝の少ない袋状の器官ができるので(文献6)、浮き袋の成立の由来には納得できる面があります。
肺の基本形は、1本か左右一対か?
「魚の肺」の研究者の間では、肺の基本形が、陸上の四足動物のように、左右1対なのか、1本の管なのか?という論争があったようです。その論点の一つは、ポリプテルスの肺が、図5の線画のように、左右1対の器官のように見えたことです。
この問題を解決すべく、最近、これらの魚類の固定標本の中の臓器の配置をX線トモグラフィー(いわゆるX―線CT画像)で解析した研究が発表されました。ただ、普通のX―線CT画像では臓器を構成する組織の微細構造がわからないので、位相差コントラストという方法を採り、そのためには強力なX線が必要であったために、日本の播磨のSpring8、フランスのSOLEIL(ソレイユ)といったシンクロトロンが使われました(文献7)。
その結果を図8に示します。それぞれの魚の断面図の左の図は前側で肺が前腸(食道)から分岐したあたり、右の図はもっと後側で、肺が広がったあたりの断面で、肺の部分は茶色に着色されています。
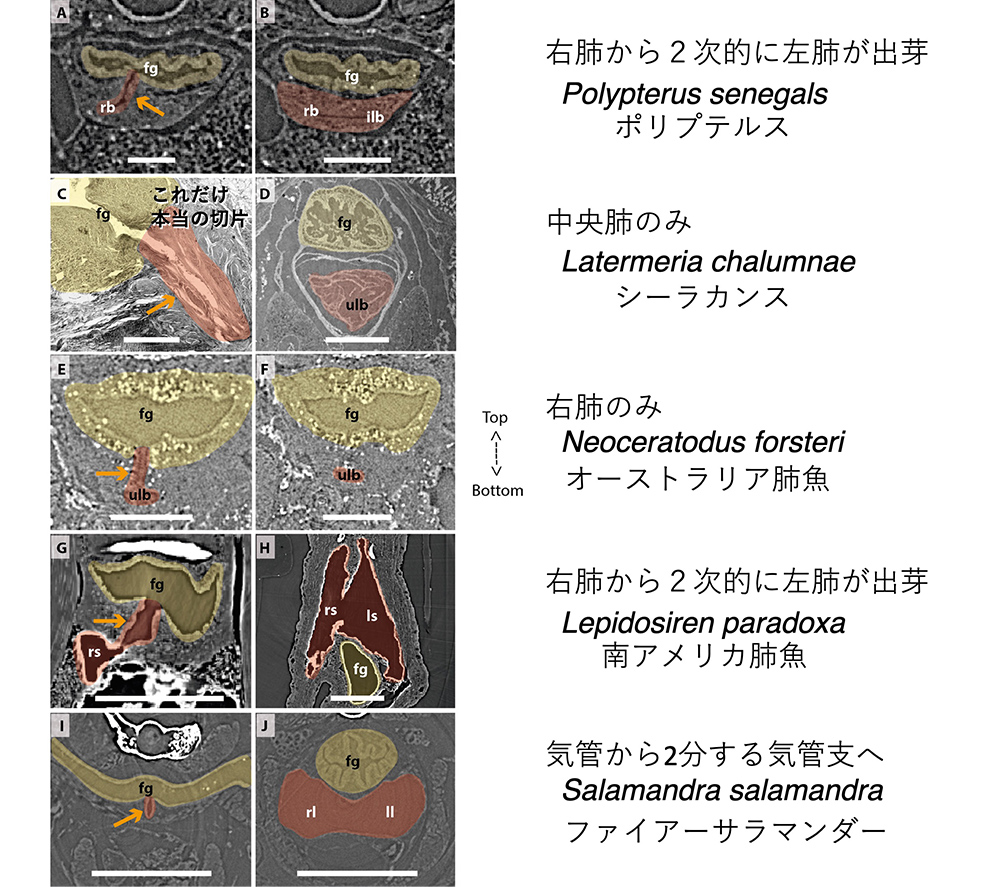
(図8)X線トモグラフィーによって描画された、各動物の肺組織の食道(前腸)とのつながり(文献7 Figure 6による)。スケールバー(白線)は以下の通り:0.25mm (A, B); 3.0mm (C); 1.0mm (D); 0.1mm (E, F); 0.5mm (G, H); 1.25mm (I, J).
この結果から、ポリプテルスは右側(向かって左側)に1本の肺の管ができ、後方で2本に見えるような広がりを作る。シーラカンスは1本の肺。オーストラリア肺魚も1本の肺、南アメリカ肺魚やアフリカ肺魚は、右肺から2次的に左肺が出芽することが明らかになりました。左右1対の肺ができるのは陸上の4足動物に限られます。結論は、肺の基本形は1本の管だということです。この結果を受けて、魚類の「肺」の模式的な線画が図9Aのように描き換えられました。
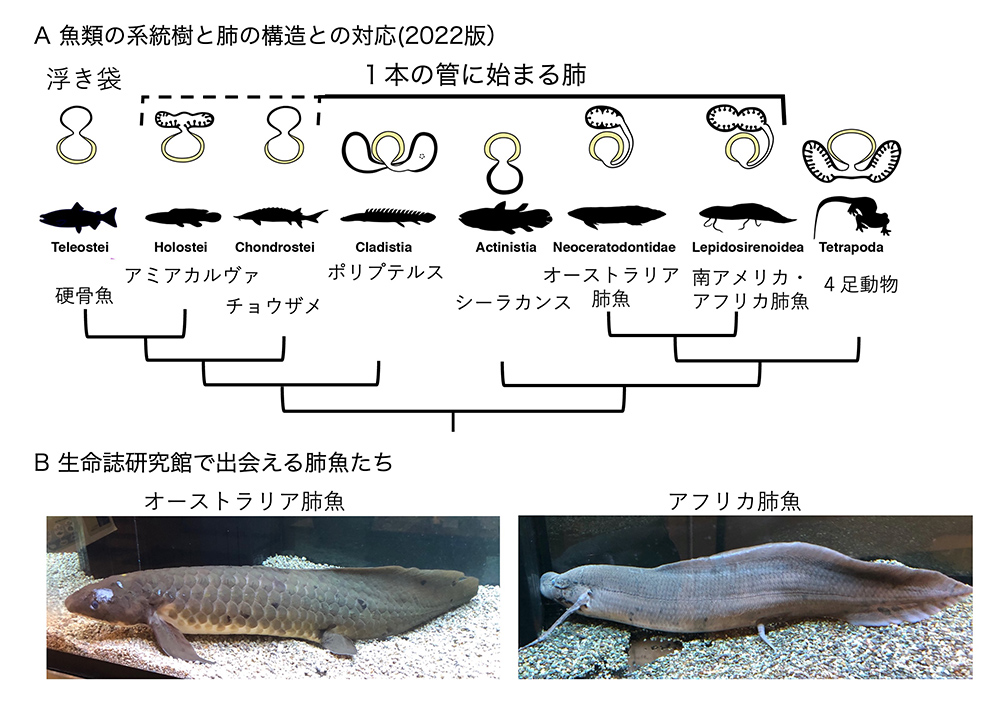
(図9)左右一対の肺を持つのは4足動物だけである:肺魚の種間に見る、肺の発達。Aは、文献7 Figure 7を改変。
生命誌研究館の入り口で、2匹の肺魚が皆様をお出迎えしています。左側の水槽にいるのはオーストラリア肺魚で、肺が未発達なので、鰓で呼吸しながら補助的に肺を使っています。右側の水槽にいるのはアフリカの肺魚で、発達した肺を持つ代わりに鰓を失い、時々「息つぎ」のために水面に上がってきます(図9B)。肺魚からさらに発達した私たちの肺の歴史を感じながら、これらの動物を鑑賞していただくのも一興かも知れません。
引用文献
[1] Teramoto M, Sugawara R, Minegishi K, Uchikawa M, Takemoto T, Kuroiwa A, Ishii Y, Kondoh H. (2020). The absence of SOX2 in the anterior foregut alters the esophagus into trachea and bronchi in both epithelial and mesenchymal components. Biol Open.9:bio048728. doi: 10.1242/bio.048728.
[2] Trisno SL, Philo KED, McCracken KW, Catá EM, Ruiz-Torres S, Rankin SA, Han L, Nasr T, Chaturvedi P, Rothenberg ME, Mandegar MA, Wells SI, Zorn AM, Wells JM. (2018). Esophageal organoids from human pluripotent stem cells delineate Sox2 functions during esophageal specification. Cell Stem Cell. 23:501-515.e7. doi: 10.1016/j.stem.2018.08.008.
[3] Kishimoto K, Furukawa KT, Luz-Madrigal A, Yamaoka A, Matsuoka C, Habu M, Alev C, Zorn AM, Morimoto M. (2020). Bidirectional Wnt signaling between endoderm and mesoderm confers tracheal identity in mouse and human cells. Nat Commun. 11:4159. doi: 10.1038/s41467-020-17969-w.
[4] Tatsumi N, Kobayashi R, Yano T, Noda M, Fujimura K, Okada N, Okabe M. (2016). Molecular developmental mechanism in polypterid fish provides insight into the origin of vertebrate lungs. Sci Rep. 6:30580. doi: 10.1038/srep30580.
[5] Uchikawa M, Ishida Y, Takemoto T, Kamachi Y, Kondoh H. (2003). Functional analysis of chicken Sox2 enhancers highlights an array of diverse regulatory elements that are conserved in mammals. Dev Cell. 4(4):509-19. doi: 10.1016/s1534-5807(03)00088-1.
[6] Arora R, Metzger RJ, Papaioannou VE. (2012). Multiple roles and interactions of Tbx4 and Tbx5 in development of the respiratory system. PLoS Genet. 8(8):e1002866. doi: 10.1371/journal.pgen.1002866.
[7] Cupello C, Hirasawa T, Tatsumi N, Yabumoto Y, Gueriau P, Isogai S, Matsumoto R, Saruwatari T, King A, Hoshino M, Uesugi K, Okabe M, Brito PM. (2022). Lung evolution in vertebrates and the water-to-land transition. Elife. 11:e77156. doi: 10.7554/eLife.77156.
サムネイルは、文献[1] Figure 1Aより
バックナンバー
VOL.4 胚のいつ、どこで、何を発生させるのかには欠かせない抑制機構
VOL.8 動物の形の多様性という名の変奏曲集I 目と顔と手